
Antagonist Temperature Variation Affects the Photosynthetic Parameters and Secondary Metabolites of Ocimum basilicum L. and Salvia officinalis L.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
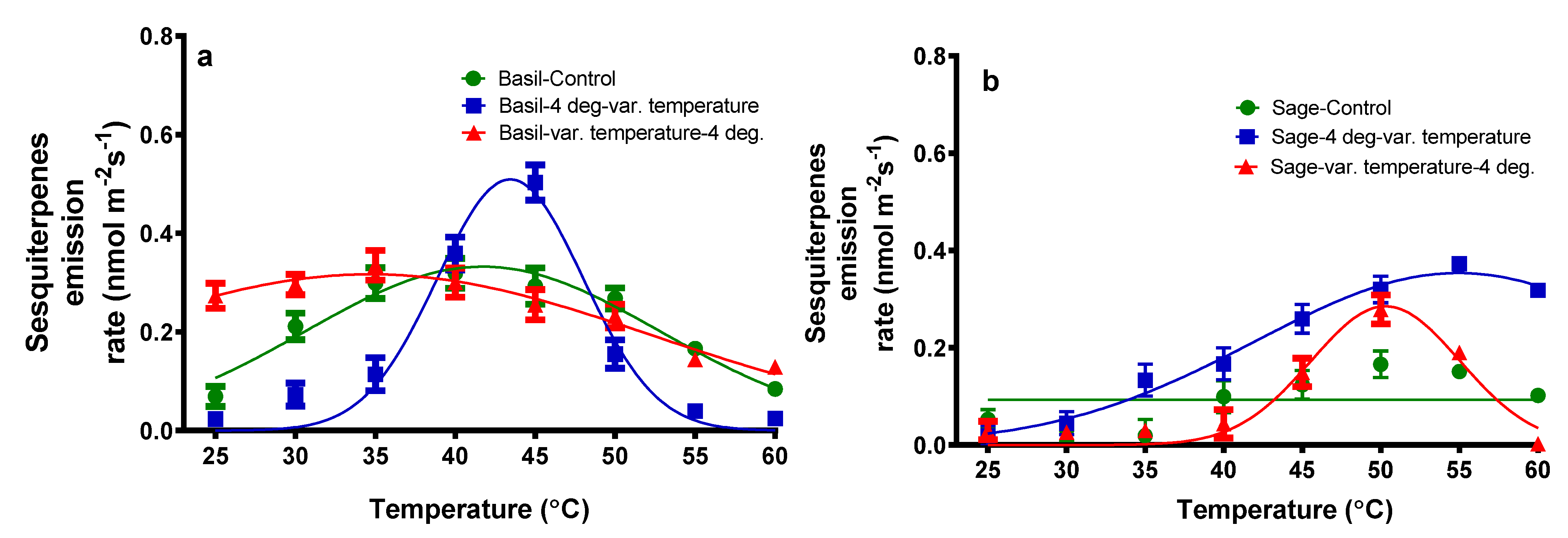{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effects of the Variation of Temperatures on Photosynthetic Characteristics
2.2. The Influence of the Variation of Temperatures on the Volatile Organic Compounds Emission
2.3. The Influence of the Variation of Temperatures on the Photosynthetic Pigments
2.4. The Influence of the Variation of Temperatures on the Total Phenolic Content—Folin−Ciocalteu Method
2.5. The Influence of the Variation of Temperatures on Total Flavonoid Content
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Photosynthetic Measurements
4.3. Volatile Sampling and GC–MS Analyses
4.4. Chromatographic Analysis of Photosynthetic Pigments
4.5. Total Phenolic Content—Folin− Ciocalteu Method
4.6. Flavonoid Content Analysis
4.7. Statistical Analysis and Data Handling
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codur, A.-M. Global Climate Change: Science and Economics. In Environmental and Natural Resource Economics, 5th ed.; Harris, J.M., Roach, B., Eds.; Routledge: New York, NY, USA, 2021; Volume 1, pp. 327–356. [Google Scholar]
- New, M.; Liverman, D.; Schroder, H.; Anderson, K. Four degrees and beyond: The potential for a global temperature increase of four degrees and its implications. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 6–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogelj, J.; Hare, W.; Lowe, J.; van Vuuren, D.P.; Riahi, K.; Matthews, B.; Hanaoka, T.; Jiang, K.; Meinshausen, M. Emission pathways consistent with a 2 °C global temperature limit. Nat. Clim. Chang. 2011, 1, 413–418. [Google Scholar] [CrossRef]
- Perkins-Kirkpatrick, S.E.; Gibson, P.B. Changes in regional heatwave characteristics as a function of increasing global temperature. Sci. Rep. 2017, 7, 12256. [Google Scholar] [CrossRef]
- Moritz, M.A.; Parisien, M.-A.; Batllori, E.; Krawchuk, M.A.; Van Dorn, J.; Ganz, D.J.; Hayhoe, K. Climate change and disruptions to global fire activity. Ecosphere 2012, 3, 1–22. [Google Scholar] [CrossRef]
- Wynne, B. Strange Weather, Again. Theory Cult. Soc. 2010, 27, 289–305. [Google Scholar] [CrossRef]
- Hughen, K.A.; Eglinton, T.I.; Xu, L.; Makou, M. Abrupt tropical vegetation response to rapid climate changes. Science 2004, 304, 1955–1959. [Google Scholar] [CrossRef] [Green Version]
- Shindell, D. Estimating the potential for twenty-first century sudden climate change. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2007, 365, 2675–2694. [Google Scholar] [CrossRef]
- Pautasso, M.; Döring, T.F.; Garbelotto, M.; Pellis, L.; Jeger, M.J. Impacts of climate change on plant diseases—Opinions and trends. Eur. J. Plant Pathol. 2012, 133, 295–313. [Google Scholar] [CrossRef] [Green Version]
- Cavaliere, C. The effects of climate change on medicinal and aromatic plants. Herb. Gram 2009, 81, 44–57. [Google Scholar]
- Copolovici, L.; Lupitu, A.; Moisa, C.; Taschina, M.; Copolovici, D.M. The Effect of Antagonist Abiotic Stress on Bioactive Compounds from Basil (Ocimum basilicum). Appl. Sci. 2021, 11, 9282. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Haverd, V.; Brendel, O.; Le Thiec, D.; Guehl, J.-M.; Cuntz, M. Robust Response of Terrestrial Plants to Rising CO2. Trends Plant Sci. 2019, 24, 578–586. [Google Scholar] [CrossRef]
- Yu, L.; Dong, H.; Huang, Z.; Korpelainen, H.; Li, C. Elevated CO2 causes different growth stimulation, water- and nitrogen-use efficiencies, and leaf ultrastructure responses in two conifer species under intra- and interspecific competition. Tree Physiol. 2021, 41, 2082–2095. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Orth, R.; Reichstein, M.; Migliavacca, M.; Zaehle, S.; Winkler, A. How does increasing CO2 influence the land-atmosphere exchange of carbon and water in response to soil and air dryness? In Proceedings of the 23rd EGU General Assembly, Online, 19–30 April 2021; p. EGU21-13494. [Google Scholar]
- Pan, Y.; Wang, Z.; Zhao, S.-W.; Wang, X.; Li, Y.-S.; Liu, J.-N.; Wang, S.; Xi, J.-H. The herbivore-induced plant volatile tetradecane enhances plant resistance to Holotrichia parallela larvae in maize roots. Pest Manag. Sci. 2022, 78, 550–560. [Google Scholar] [CrossRef] [PubMed]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, E.F.; Pallini, A.; Janssen, A. Herbivore performance and plant defense after sequential attacks by inducing and suppressing herbivores. Insect Sci. 2019, 26, 108–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Copolovici, L.; Pag, A.; Kännaste, A.; Bodescu, A.; Tomescu, D.; Copolovici, D.; Soran, M.-L.; Niinemets, Ü. Disproportionate photosynthetic decline and inverse relationship between constitutive and induced volatile emissions upon feeding of Quercus robur leaves by large larvae of gypsy moth (Lymantria dispar). Environ. Exp. Bot. 2017, 138, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Loc, M.; Milošević, D.; Ignjatov, M.; Ivanović, Ž.; Budakov, D.; Grahovac, J.; Vlajkov, V.; Pajčin, I.; Grahovac, M. First Report of Pectobacterium punjabense causing potato soft rot and blackleg in Serbia. Plant Dis. 2022, 106, 1513. [Google Scholar] [CrossRef]
- Marr, A.; Casonato, S.; Jones, E. Short Communication: Latent Detection of Downy Mildew (Peronospora pisi) in Bioassays against Pisum sativum. Pathogens 2021, 10, 1312. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.R.; Cockshull, K.E.; Cave, C.R.J. Effect of Temperature on the Growth and Development of Tomato Fruits. Ann. Bot. 2001, 88, 869–877. [Google Scholar] [CrossRef]
- Karlsson, M.; Werner, J. Temperature affects leaf unfolding rate and flowering of cyclamen. HortScience 2001, 36, 292–294. [Google Scholar] [CrossRef]
- Knight, M.R.; Knight, H. Low-temperature perception leading to gene expression and cold tolerance in higher plants. New Phytol. 2012, 195, 737–751. [Google Scholar] [CrossRef]
- Niinemets, Ü. When leaves go over the thermal edge. Plant Cell Environ. 2018, 41, 1247–1250. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Hafeez, M. Thermotolerance of pearl millet and maize at early growth stages: Growth and nutrient relations. Biol. Plant. 2004, 48, 81–86. [Google Scholar] [CrossRef]
- Ramin, A.A. Effects of salinity and temperature on germination and seedling establishment of sweet basil (Ocimum basilicum L.). J. Herbs Spices Med. Plants 2006, 11, 81–90. [Google Scholar] [CrossRef]
- Akbari, G.A.; Soltani, E.; Binesh, S.; Amini, F. Cold tolerance, productivity and phytochemical diversity in sweet basil (Ocimum basilicum L.) accessions. Ind. Crops Prod. 2018, 124, 677–684. [Google Scholar] [CrossRef]
- Lin, K.-H.; Lin, T.-Y.; Wu, C.-W.; Chang, Y.-S. Protective Effects of Salicylic Acid and Calcium Chloride on Sage Plants (Salvia officinalis L. and Salvia elegans Vahl) under High-Temperature Stress. Plants 2021, 10, 2110. [Google Scholar] [CrossRef]
- Kim, K.; Portis, A.R., Jr. Temperature dependence of photosynthesis in Arabidopsis plants with modifications in Rubisco activase and membrane fluidity. Plant Cell Physiol. 2005, 46, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Hüve, K.; Bichele, I.; Tobias, M.; Niinemets, Ü. Heat sensitivity of photosynthetic electron transport varies during the day due to changes in sugars and osmotic potential. Plant Cell Environ. 2006, 29, 212–228. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stress, Volume 1: Chilling, Freezing, and High Temperature Stresses; Academic Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Filella, I.; Wilkinson, M.J.; Llusia, J.; Hewitt, C.N.; Peñuelas, J. Volatile organic compounds emissions in Norway spruce (Picea abies) in response to temperature changes. Physiol. Plant. 2007, 130, 58–66. [Google Scholar] [CrossRef]
- Loreto, F.; Barta, C.; Brilli, F.; Nogues, I. On the induction of volatile organic compound emissions by plants as consequence of wounding or fluctuations of light and temperature. Plant Cell Environ. 2006, 29, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Hartikainen, K.; Riikonen, J.; Nerg, A.-M.; Kivimäenpää, M.; Ahonen, V.; Tervahauta, A.; Kärenlampi, S.; Mäenpää, M.; Rousi, M.; Kontunen-Soppela, S.; et al. Impact of elevated temperature and ozone on the emission of volatile organic compounds and gas exchange of silver birch (Betula pendula Roth). Environ. Exp. Bot. 2012, 84, 33–43. [Google Scholar] [CrossRef]
- Grote, R.; Monson, R.K.; Niinemets, Ü. Leaf-level models of constitutive and stress-driven volatile organic compound emissions. In Biology, Controls and Models of Tree Volatile Organic Compound Emissions; Springer: Berlin/Heidelberg, Germany, 2013; pp. 315–355. [Google Scholar]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, Ü. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Fares, S.; Oksanen, E.; Lännenpää, M.; Julkunen-Tiitto, R.; Loreto, F. Volatile emissions and phenolic compound concentrations along a vertical profile of Populus nigra leaves exposed to realistic ozone concentrations. Photosynth. Res. 2010, 104, 61–74. [Google Scholar] [CrossRef]
- Wu, G.; Johnson, S.K.; Bornman, J.F.; Bennett, S.J.; Clarke, M.W.; Singh, V.; Fang, Z. Growth temperature and genotype both play important roles in sorghum grain phenolic composition. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Wang, S.Y.; Zheng, W. Effect of plant growth temperature on antioxidant capacity in strawberry. J. Agric. Food Chem. 2001, 49, 4977–4982. [Google Scholar] [CrossRef]
- Hendawy, S.F.; Khalid, K.A. Response of sage (Salvia officinalis L.) plants to zinc application under different salinity levels. J. Appl. Sci. Res 2005, 1, 147–155. [Google Scholar]
- Duerbeck, K. EU Market Survey 2004. Natural Ingredients for Pharmaceuticals. 2004. Available online: http://repositorio.promperu.gob.pe/bitstream/handle/123456789/1430/EU_market_survey_2004_natural_ingredients_pharmaceuticals_2004_keyword_principal.pdf?sequence=1 (accessed on 19 October 2021).
- Chang, X.; Alderson, P.; Wright, C. Effect of temperature integration on the growth and volatile oil content of basil (Ocimum basilicum L.). J. Hortic. Sci. Biotechnol. 2005, 80, 593–598. [Google Scholar] [CrossRef]
- Behnke, K.; Ghirardo, A.; Janz, D.; Kanawati, B.; Esperschütz, J.; Zimmer, I.; Schmitt-Kopplin, P.; Niinemets, Ü.; Polle, A.; Schnitzler, J.P.; et al. Isoprene function in two contrasting poplars under salt and sunflecks. Tree Physiol 2013, 33, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitao, M.; Kitaoka, S.; Harayama, H.; Tobita, H.; Agathokleous, E.; Utsugi, H. Canopy nitrogen distribution is optimized to prevent photoinhibition throughout the canopy during sun flecks. Sci. Rep. 2018, 8, 503. [Google Scholar] [CrossRef] [PubMed]
- Hemantaranjan, A.; Bhanu, A.N.; Singh, M.; Yadav, D.; Patel, P.; Singh, R.; Katiyar, D.J.A.P.A.R. Heat stress responses and thermotolerance. Adv. Plants Agric. Res. 2014, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Sánchez, E.; Ruiz, J.M.; Romero, L. Influence of temperature on biomass, iron metabolism and some related bioindicators in tomato and watermelon plants. Plant Physiol. 2003, 160, 1065–1071. [Google Scholar] [CrossRef] [Green Version]
- Murkowski, A. Heat Stress and Spermidine: Effect on Chlorophyll Fluorescence in Tomato Plants. Biol. Plant. 2001, 44, 53–57. [Google Scholar] [CrossRef]
- Rainwater, D.T.; Gossett, D.R.; Millhollon, E.P.; Hanna, H.Y.; Banks, S.W.; Lucas, M.C. The relationship between yield and the antioxidant defense system in tomatoes grown under heat stress. Free Radic. Res. 1996, 25, 421–435. [Google Scholar] [CrossRef]
- Chumyam, A.; Whangchai, K.; Jungklang, J.; Faiyue, B.; Saengnil, K.J.S.A. Effects of heat treatments on antioxidant capacity and total phenolic content of four cultivars of purple skin eggplants. Sci. Asia 2013, 39, 246–251. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, A. Cold Stress and Photosynthesis. In Photosynthesis, Productivity and Environmental Stress; John Wiley & Sons: Hoboken, NJ, USA, 2019; pp. 27–37. [Google Scholar]
- Beck, E.H.; Heim, R.; Hansen, J. Plant resistance to cold stress: Mechanisms and environmental signals triggering frost hardening and dehardening. J. Biosci. 2004, 29, 449–459. [Google Scholar] [CrossRef]
- Zhu, J.; Liang, Y.; Ding, Y.; Li, Z. Effect of silicon on photosynthesis and its related physiological parameters in two winter wheat cultivars under cold stress. Zhongguo Nong Ye Ke Xue 2006, 39, 1780–1788. [Google Scholar]
- Zhang, R.; Sharkey, T.D. Photosynthetic electron transport and proton flux under moderate heat stress. Photosynth. Res. 2009, 100, 29–43. [Google Scholar] [CrossRef]
- Copolovici, L.O.; Filella, I.; Llusià, J.; Niinemets, U.; Peñuelas, J. The capacity for thermal protection of photosynthetic electron transport varies for different monoterpenes in Quercus ilex. Plant Physiol. 2005, 139, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guenther, A.B.; Zimmerman, P.R.; Harley, P.C.; Monson, R.K.; Fall, R. Isoprene and monoterpene emission rate variability: Model evaluations and sensitivity analyses. J. Geophys. Res. Atmos. 1993, 98, 12609–12617. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Ye, J.; Li, S.; Niinemets, Ü. Regulation of Floral Terpenoid Emission and Biosynthesis in Sweet Basil (Ocimum basilicum). J. Plant Growth Regul. 2016, 35, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Kalisz, A.; Jezdinský, A.; Pokluda, R.; Sękara, A.; Grabowska, A.; Gil, J. Impacts of chilling on photosynthesis and chlorophyll pigment content in juvenile basil cultivars. Hortic. Environ. Biotechnol. 2016, 57, 330–339. [Google Scholar] [CrossRef]
- Kännaste, A.; Copolovici, L.; Niinemets, Ü. Gas chromatography–mass spectrometry method for determination of biogenic volatile organic compounds emitted by plants. In Plant Isoprenoids; Springer: Berlin/Heidelberg, Germany, 2014; pp. 161–169. [Google Scholar]
- Opriş, O.; Copaciu, F.; Soran, M.L.; Ristoiu, D.; Niinemets, Ü.; Copolovici, L. Influence of nine antibiotics on key secondary metabolites and physiological characteristics in Triticum aestivum: Leaf volatiles as a promising new tool to assess toxicity. Ecotoxicol. Environ. Saf. 2013, 87, 70–79. [Google Scholar] [CrossRef]
- Pag, A.I.; Radu, D.G.; Popa, M.I.; Sirghie, C. Flaxseed cake—A sustainable source of antioxidant and antibacterial extracts. Cellul. Chem. Technol. 2014, 48, 265–273. [Google Scholar]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Copolovici, L.; Copolovici, D.M.; Moisa, C.; Lupitu, A. Antagonist Temperature Variation Affects the Photosynthetic Parameters and Secondary Metabolites of Ocimum basilicum L. and Salvia officinalis L. Plants 2022, 11, 1806. https://doi.org/10.3390/plants11141806
Copolovici L, Copolovici DM, Moisa C, Lupitu A. Antagonist Temperature Variation Affects the Photosynthetic Parameters and Secondary Metabolites of Ocimum basilicum L. and Salvia officinalis L. Plants. 2022; 11(14):1806. https://doi.org/10.3390/plants11141806
Chicago/Turabian StyleCopolovici, Lucian, Dana M. Copolovici, Cristian Moisa, and Andreea Lupitu. 2022. "Antagonist Temperature Variation Affects the Photosynthetic Parameters and Secondary Metabolites of Ocimum basilicum L. and Salvia officinalis L." Plants 11, no. 14: 1806. https://doi.org/10.3390/plants11141806