
Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species


1
Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México, Mexico City 04510, Mexico
2
Departamento de Ciencias Ambientales y del Suelo, Instituto de Geología, Universidad Nacional Autónoma de México, Mexico City 04510, Mexico
*
Author to whom correspondence should be addressed.
Plants 2022, 11(12), 1554; https://doi.org/10.3390/plants11121554
Submission received: 21 April 2022
/
Revised: 28 May 2022
/
Accepted: 9 June 2022
/
Published: 12 June 2022
(This article belongs to the Special Issue Advance in Orchidaceae Research)
Abstract
:In the valuable orchid genus Cypripedium, the section Irapeana consists of a distinctive group of Mesoamerican species that is formed by Cypripedium dickinsonianum Hágsater, C. irapeanum Lex., and C. molle Lindl. All lady slipper orchids exhibit different distributions and abundances. Data analysis that used herbarium accessions and field investigations indicated that the habitats of these three species have been dramatically reduced. Prospecting for suitable habitats based on climatic, vegetation, and soil parameters allows us to predict potential distributions. Conservation strategies, such as ex situ propagation by asymbiotic and symbiotic approaches, have indicated that the culture media used are a determining factor for seedling development. Mycorrhizal isolates play a main role in the compatibility and further development of germinated seeds. The fungi isolated from adult plants belong to two different families, which makes it possible that widely distributed C. irapeanum populations will be fungal-specific as well as restricted for C. molle. Root mycorrhization patterns occur high on the secondary roots. In contrast with other species of the genus, in situ germination can occur over a short period of two months, but we have documented periods as long as ten years. Cypripedium is a highly problematic genus for ex situ conservation because the germination requirements and cultures are poorly documented, and there is great urgency for in situ conservation to develop strategies for identifying hotspot habitats and actualize the protection status to avoid extinction of this genus.
1. Introduction
The terrestrial genus Cypripedium is one of the most appreciated in the orchid family and is found in the natural environment, botanical gardens, natural parks, and scientific and private collections [1,2]. The origin of the name comes from the Greek root Cypris that refers to Aphrodite’s sandal pedilon because of the globose-sac-shaped flower lips. Along with modifications of sepals and petals, columns with two anthers and the presence of staminodes make them unique relative to all other orchids. The genus contains approximately 50 species that are distributed in the Northern Hemisphere in mountain woodlands, grasslands, shrubs, or swamps, and is mainly associated with Quercus or Pinus forests [2,3,4,5,6,7,8,9]. All species are endangered by overcollection, anthropogenic activities, and climate change [10,11].
Mesoamerica has been postulated to be the origin center of the genus Cypripedium with the section Irapeana, which is a sister clade to the other members of this taxa [2,4,7,12,13,14]. The section contains three species, namely, C. dickinsonianum Hágsater, C. irapeanum Lex., and C. molle Lindl. [8]. C. irapeanum from Irapeo, which is located in Michoacán State of Mexico, is the type species, but the type locality no longer exists [13,14,15]. There is remarkable morphological variability among C. irapeanum populations, which are likely to consist of a complex of species to be resolved by using molecular markers [16]. Morphological variations, mainly in size, are also observed in C. dickinsonianum populations from the northeastern Sierra Madre compared to those in the Chiapas and Guatemala Sierras. Mesoamerican Cypripedium flowers show remarkable morphological characteristics: the largest flowers are found for taller C. irapeanum plants, and the smallest flowers are found for C. dickinsonianum (Figure 1a–c). The colors vary from a striking pale-yellow to canary yellow, and the staminode (sterile stamen) is often showy to welcome the insect, which makes its way to a backdoor exit into the pouch. The dorsal sepals are generally not as wide as those in the rest of the cypripediums, and the petals are wider and have slightly smaller synsepals. C. irapeanum and C. molle also show distinctive sepia-reddish staining inside the lobes, and they have small transparent “windows” over the lip surfaces. The local name of the genus is “pichohuaxtle”, derived from the Náhuatl dialect, which means “bulls’ eggs” [17]. Mexican children crush the showy inflated lip of the flowers for use as whistles or balloons [4,14,18].
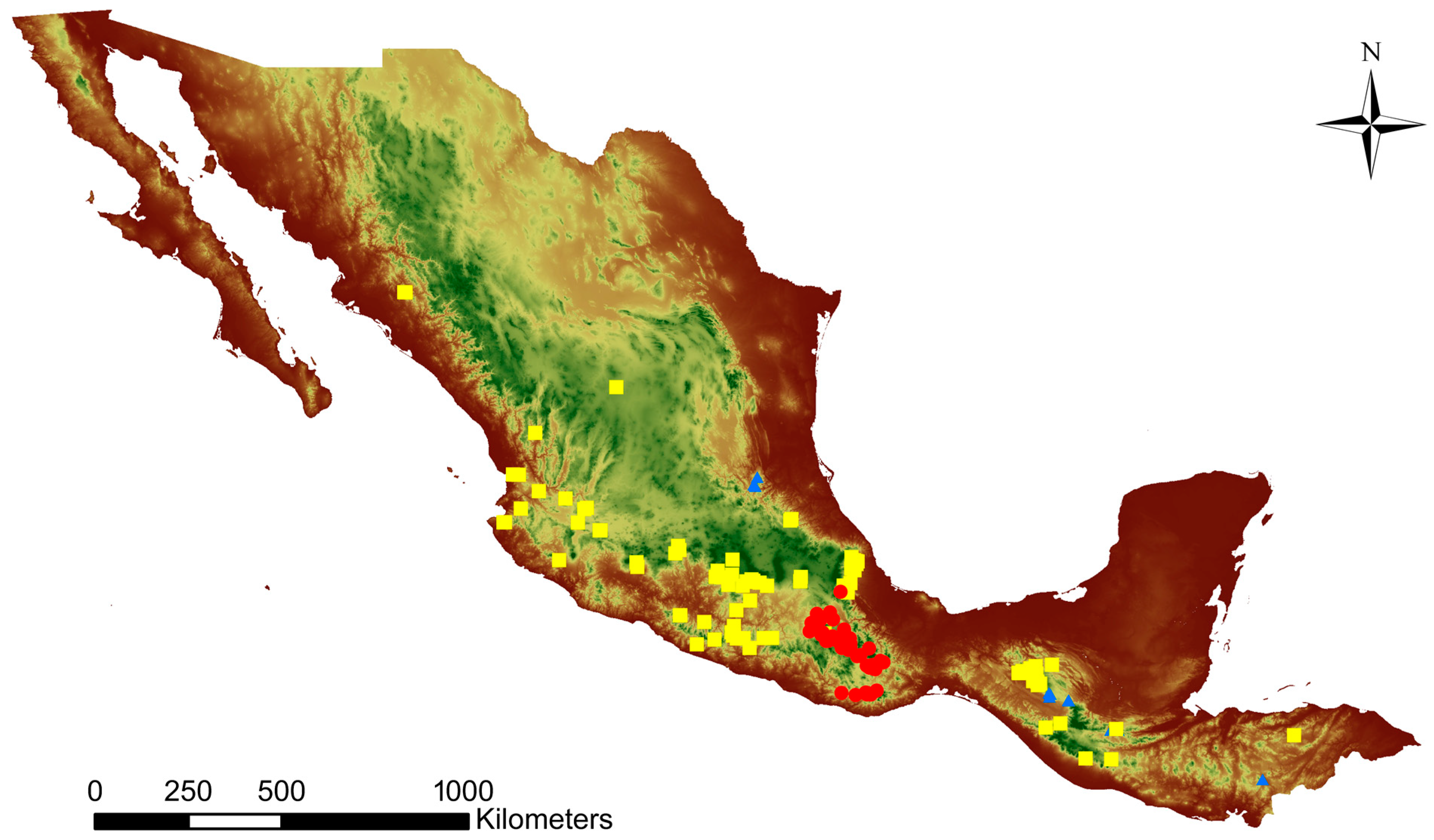
The Mesoamerican species have dissimilar distributions. C. dickinsonianum is restricted to small populations in the Mexican states of Queretaro and Chiapas and in Guatemala and grows on light slopes with Juniperus, and is sometimes sympatric with C. irapeanum [13,14,18]. In contrast, C. irapeanum is widely distributed in shrubs and grasslands or is associated with Quercus and Pinus in several Mexican states (e.g., Chiapas, Puebla, Mexico, Morelos, Michoacán, Guerrero, Colima, Nayarit, Sinaloa, and Veracruz), and is found in some locations in Guatemala. Cypripedium molle is restricted to the Mexican state of Oaxaca and grows in similar forest associations as C. irapeanum, but with a difference wherein it develops close to disturbed sites such as roadsides [19] (Figure 2).
2. Mesoamerican Slipper Orchids: Unique and Critically Endangered
Many factors increase the risk of loss of these species and developing strategies for their conservation is extremely urgent. International treatment CITES (Convention on International Trade in Endangered Species of Wild Fauna and Flora) considers C. dickinsonianum as endangered (EN), C. irapeanum as vulnerable (VU), and C. molle as near threatened (NT) [20]. Mexican policies recognize C. dickinsonianum as protected and C. irapeanum as EN, while the status of C. molle is unconsidered [21]. However, the actual endangered status of all Mesoamerican species is more serious: by using georeferenced entries from Mexican National herbarium records from 1954 to 2013 (176 entries) (Salazar-Chávez, G., personal database), data from previous papers [18], and confirmed records from the website inaturalist.org [22], we obtained 11 records for C. dickinsonianum, 102 for C. irapeanum, and 57 records for C. molle. We determined by using geographic information system techniques, satellite images, georeferenced databases [23], and ecological niche modeling projections [24] that only 3, 43, and 27 of the records, respectively, maintained their original habitats, with a loss of 58% for both C. dickinsonianum and C. irapeanum and 53% for C. molle (Figure 3). Most of the remnants confirmed that populations are endangered due to their proximity to urban sites or sites with probable habitat transformation: from field investigations conducted in 2018 and 2019, we visited 27 recorded C. molle populations, and only 3 of them could be found, while the others had been destroyed at the end of 2019 by Agave angustifolia plantations used for mezcal production, an alcoholic drink, whose consumption has popularized since the Appellation of Origin was obtained by only the Mexican state of Oaxaca [25]. In the case of C. irapeanum, two of five studied populations were subjected to pillage of flowers and plants. Additional threats come from habitat transformation due to road expansion and the establishment of illegal trash dumps. C. dickinsonianum is the most vulnerable, and the remaining four populations, which represent half of the total number of records, are near highways and urban centers.
Due to the swift loss of natural habitats for these Mesoamerican species, identification of potential habitats was achieved to identify possible new locations to find or reintroduce populations in the future. Some climatic factors may strongly determine their distributions [18], so ecological preferences and ecological niche modeling have been studied by using the WorldClim-Global Climate Data, which include the temperature seasonality, mean annual precipitation, annual temperature range, mean temperature in the coldest quarter, annual mean precipitation, and precipitation seasonality [26]. We obtained hypothetical projections for the three species by using the maximum entropy algorithm calculated with MAXENT software [27]. To estimate the model, we used herbarium and online records obtained from the website inaturalist.org [22], climatic variables, soil conditions’ layers (e.g., total carbon and nitrogen, bulk density, water-holding capacity, moisture content, and wilting point) [28], altitudinal records, and vegetation types [23] (Table S1), and these data had previously been transformed to compatible formats in ArcGIS© [29].
The resulting projections of the potential habitats for each species indicate that C. irapeanum and C. dickinsonianum are sympatric in some habitats, whereas certain potential distribution habitats have not yet been recorded (Figure 4a,b). For C. molle, the estimated distribution is restricted to the gap located between the union of the eastern and south Sierra Madre located in the physiographic province of Oaxaca´s Sierras Centrales (Figure 4c). The inclusion of soil conditions generates more robust models for suitable habitat projections; however, it is not clear whether soil conditions have a direct impact on plant requirements or the associated microorganisms, such as orchid mycorrhizal fungi (OMF), as we conduct further research [30].
3. Underground Growth Pattern in Different Habitats
We analyzed the underground rhizome growth after conducting the field investigations for three habitats: two for C. irapeanum (Quercus forest in the states of Mexico, Morelos, and Puebla, and Tropical Deciduous Forest (TDF) in Veracruz State) and one for C. molle (Quercus forest in Oaxaca State). Both species show consistent patterns of underground root growth that are generally less than 10 cm-deep and are located between an undecomposed litter layer and a layer with high organic matter content that is very similar to the growth of other Cypripedium species [4,31] (Figure 5). Mesoamerican species develop short rhizomes and an annual stem with several large roots that can persist for years by storing carbohydrates in the form of starch. For other species in the genus, the root systems may live for nearly 14 years, which allows these plants to remain dormant for long periods without aerial shots until adequate conditions for vegetative growth are met. Many years (e.g., ca. 7–16) are necessary to develop full-grown flowering plants from seedlings of lady slipper orchid species [4,32,33,34]. For C. irapeanum, the populations in Quercus forest grow under layers rich in organic matter on volcanic-derived acidic soils (pH of 5.6), while populations in TDF grow in poor soils, with high levels of calcium, little organic matter, and a pH of approximately 7.5 [14,35]. Both populations develop under seasonally dry environments. It is worth mentioning that in five years of study on these populations, only one germination event was recorded for each population (Figure 6a,b). The incidence of in situ germination has been estimated to be extremely low (e.g., approximately 0.001%) [8] and can occur over a period of two months (Figure 6c–e) or ten years after seed dispersion (Figure 6f–h). It is not clear whether these scarce germination events depend on the potential of a habitat to provide a germination niche (for example, until colonization by compatible symbiotic fungi) or on the natural long-term dormancy of seeds as a survival mechanism. OMFs are needed for germination and carbon acquisition throughout life to support survival during dormant states, quite common in the genus Cypripedium [4,32,36,37,38,39]. In general, the in situ germination requirements for this genus are poorly understood, and long-term studies are needed [40]. Both C. irapeanum and C. molle grow on different soils, such as reddish, clayey, lateritic, limestone soils or volcanic-derived soils, and this indicates that a broad range of diverse conditions can meet the requirements for germination and increases the potential habitats and probability of establishing new populations [41]. The genus is considered highly dependent on mycorrhizal fungi, and this could be the reason why asymbiotic propagation methods have not been completely established for many species [42,43,44].
4. Mycorrhiza Studies for Conservation Purposes
Cypripedium species, like other orchids, have two main symbiotic relationships: with insects for pollination and seed production and with OMF for germination and nutrient transfer. For pollination, there are several genera of insects that can function as Cypripedium pollinators, and despite the different reports on these insects, few actual successful pollinia removal events have been recorded [45,46,47]. In the case of Mesoamerican C. irapeanum and C. molle, Halictideae wasps have been considered as pollinators for both, and the only pollinator for C. irapeanum is considered as Lasioglossum nyctere, while C. dickinsonianum has been recognized as self-pollinating [13,14].
However, mycorrhizal symbiotic associations have more effectively studied for several species in adult plants, and scarce and erratic patterns of root colonization [37,48]. We confirm two main patterns of mycorrhiza development for Mesoamerican C. irapeanum and C. molle: On the main roots, colonization is scarce and can remain for years with highly degraded hyphal coils and large numbers of starch granules [37,48,49]. In contrast, the secondary roots, which are usually short and numerous (ca. 2–3 cm), lack starch reserves because they are actively growing and are highly colonized by hyphal coils in diverse stages of digestion, while most of them are undigested (Figure 7). Seedlings that develop by in situ germination of both species are always colonized with the same pattern as the secondary roots.
There have only been three successful attempts to isolate OMF on Cypripedium, for C. macranthos var. rebunense [43] and Mesoamerican C. irapeanum [35] and C. molle (Moreno-Camarena and Ortega-Larrocea, submitted). The mycorrhizal endophytes that were obtained from the roots of adult C. irapeanum plants from two habitats belong to the anamorphic genus Epulorhiza (hyphae less than 4 µm, pearly monilioid cells, creamy submerged colonies on PDA, and slow growth rates ca. 0.2 mm per day). Endophytes that were obtained from C. molle show characteristics of the anamorph Ceratorhiza (hyphae of more than 4 µm, barrel-shaped monilioid cells, brownish colonies on PDA, aerial mycelium, and growth rates of 0.5 mm/day) (Figure 8, Table 1) [50]. The isolation and long-term conservation of these isolates was achieved only on Green Pea Agar medium [51], AWA (Acidic Water Agar), and FIM (Fungal Isolation Medium) [52]. In media such as PDA (potato dextrose agar) or OMA (oatmeal agar), which are usually used for OMF cultivation, no long-term growth was achieved, and the isolates lost viability in both cases.
Since OMF isolation has been difficult for Cypripedium species, using molecular tools to identify the relevant fungi is usually conducted by using the fungal coils of adult roots. The genomic regions studied are the ITS (internal transcribed spacer), LSU (large subunit), and SSU (short subunit) [53,54,55]. Molecular identifications indicate that the genera are sometimes associated with distant phylogenetic fungal groups (Table S2) [56]. In addition, some of the identified fungal partners may be incidental inhabitants, pathogens, or temporal successors [36]. The most common OMFs associated with Cypripedium belong to the family Tulasnellaceae: Tulasnella cystidiophora, T. calospora, and T. deliquescens, which were found in 28, 12, and 5 species, respectively. The family Ceratobasidiaceae (Ceratobasidium cornigerum) has also been identified in C. californicum [37,44,48,57]. However, most of these identifications have been conducted on adult plants, since symbiotic germination is poorly documented and the identification of fungal germination promoters is needed [42,43,58].
To analyze the relationships among fungi that were isolated from the adult roots of C. irapeanum (Quercus forest in Mexico State and TDF in Veracruz State) and plantlets of C. molle (Quercus forest in Oaxaca State), we amplified their ITS regions by using the primer combination ITS 1/ITS 4 [36,37,48,51,57,59], and the resulting sequences were assembled and edited with Geneious (2021.0.3). The sequences are deposited in the GenBank-NCBI database. A search for the most similar sequences was conducted by using the BLAST algorithm [60,61], and alignments were conducted using MAFFT [62]. Both algorithms are contained in the Geneious software. Phylogenetic reconstruction was carried out using PHYML plugin [60,63] by using the maximum likelihood method and the Tamura-Nei model with bootstrap support of 1000 replicates (Figure 9).
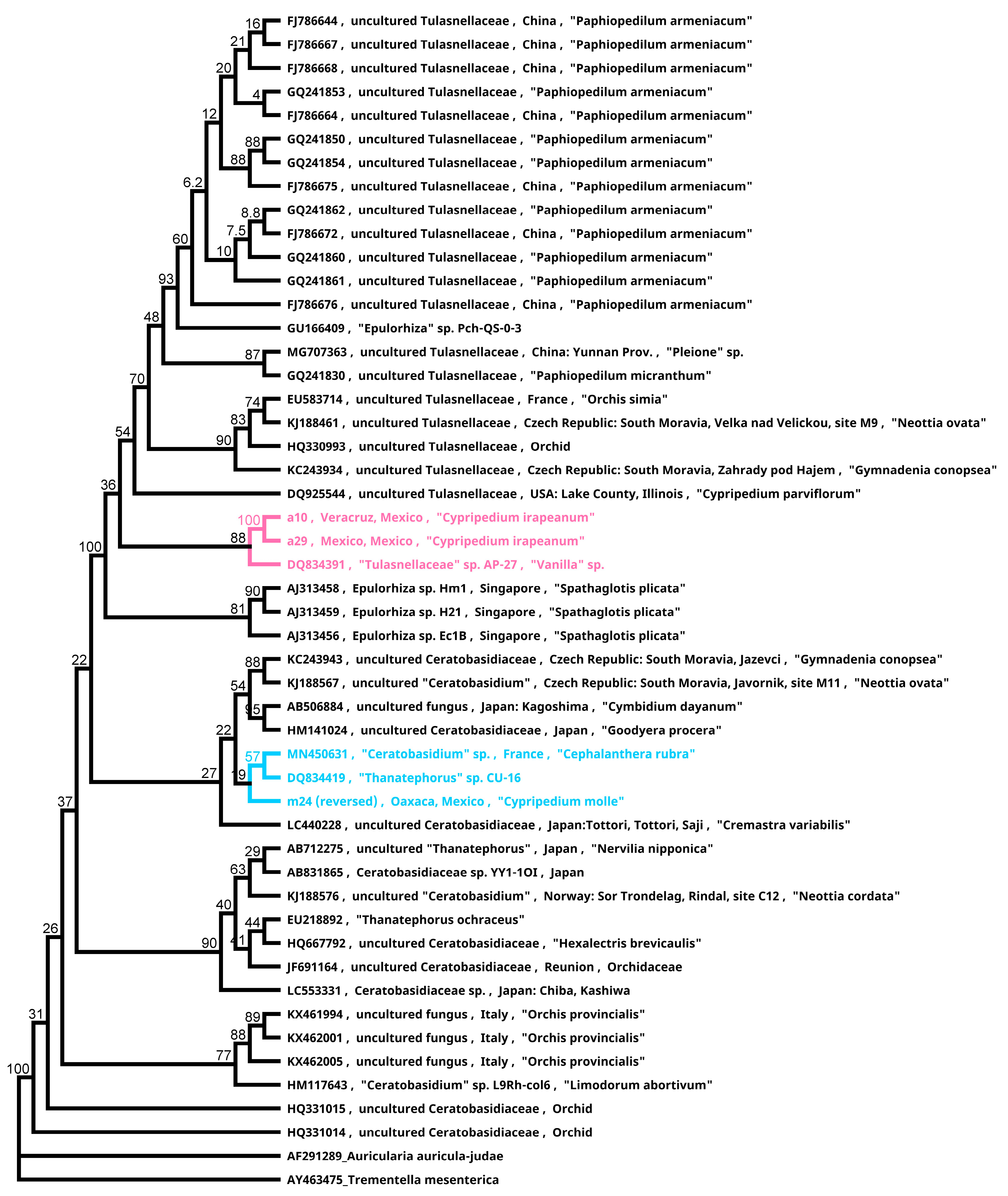
Significant results were observed from the phylogenetic reconstruction. The isolates from C. molle plantlets belong to Ceratobasidiaceae in a clade that consists of terrestrial endophytes of Vanilla spp. from Puerto Rico and Cephalanthera rubra from France [64] (Figure 9). Conversely, the isolates of the widespread C. irapeanum were recovered from different habitats (e.g., Quercus forest and TDF) and belong to Tulasnellaceae in a clade that includes a terrestrial endophyte from Vanilla [50], which indicates a probable high level of specificity to a widely distributed soil generalist fungal clade (Figure 9). This behavior is opposite to that of the North American C. californicum, a soil specialist (always grows on serpentine soils) but is associated with various species of the genus Tulasnella [48]. The same is true for the terrestrial Dichromanthus that form specific associations with a particular mycorrhizal endophyte when the species are widely distributed in several habitats and soil conditions. However, in one restricted habitat, the species were associated with several clades of the same fungal family (López-Reyes and Ortega-Larrocea, personal communication).
The fact that plantlets of C. molle associate with Ceratobasidiaceae may indicate that the fungi from this family are germination promoters, as was proposed for C. calceolus [65]. This could explain why endophytes isolated from adult plants do not promote good germination, as has been observed in C. irapeanum [35] and previously in C. macranthos var. rebunense [51]. The isolation and identification of fungi from other populations and plant stages together with cultivation by asymbiotic and symbiotic methods would provide a better understanding of germination, specificity, fungal succession during plant lifetimes, distribution of populations, and microhabitat requirements to develop conservation strategies for these unique species.
5. Symbiotic and Asymbiotic Germination and Development
Seeds of Cypripedium spp. have tests that consist of a hydrophobic double-layer rich in lignin that allows them to float on water [66,67]. At maturity, these seeds have high concentrations of abscisic acid (ABA) that can delay germination [68,69] which are probably involved in long-term survival through the years and persist for at least four years to form seed banks in soils [4,42,70]. In situ germination relies on mycorrhizal fungi that can penetrate the testa and induce germination [42,43]. Symbiotic germination under in situ conditions is poorly studied in Cypripedium and probably occurs in spring or early summer at 2–5 cm soil depths in moisture-stable sites within a pH range from 5.3 to 8.1 [31]. In some species of the genus, germination is stimulated by cold (e.g., C. calceolus, C. lentiginosum, C. macranthos var. rebunense) because they grow in temperate regions [42,43,68,71].
In vitro germination of Mesoamerican species has been studied using symbiotic and asymbiotic approaches. Asymbiotic germination was tested on PhytamaxTM (SIGMA) with sucrose 2% [72], Murashige and Skoog [73], Norstog [74], and oatmeal agar [52] at pH 5.6 after the seeds were stimulated by cold storage (4 °C) for four months to break dormancy (Moreno-Camarena and Ortega-Larrocea, in process). However, seedlings developed only on Norstog medium (Figure 10). It is worth mentioning that while Asian or European Cypripedium species germinate at 4 to 54 months after sowing [42,58], C. irapeanum can germinate at 12–14 days after sowing (das) [35]. Germination development, in vitro or in situ, begins when the testa splits and the embryo swells, which gives rise to a protocorm with a promeristematic zone (Figure 10a–c). From this, the apex begins to grow, which is followed by root formation on the opposite point of the protocorm (Figure 10d–f). The protocorms are greenish and usually have long rhizoids [31,42], except for C. irapeanum, which does not develop rhizoids under asymbiotic in vitro conditions (Figure 10d–g). Roots emerge after shoot differentiation of two foliar sheets (Figure 10g). This root system continues to grow faster over the shoot leaves and develops into several small sheets and lateral shoots (Figure 10h–k).
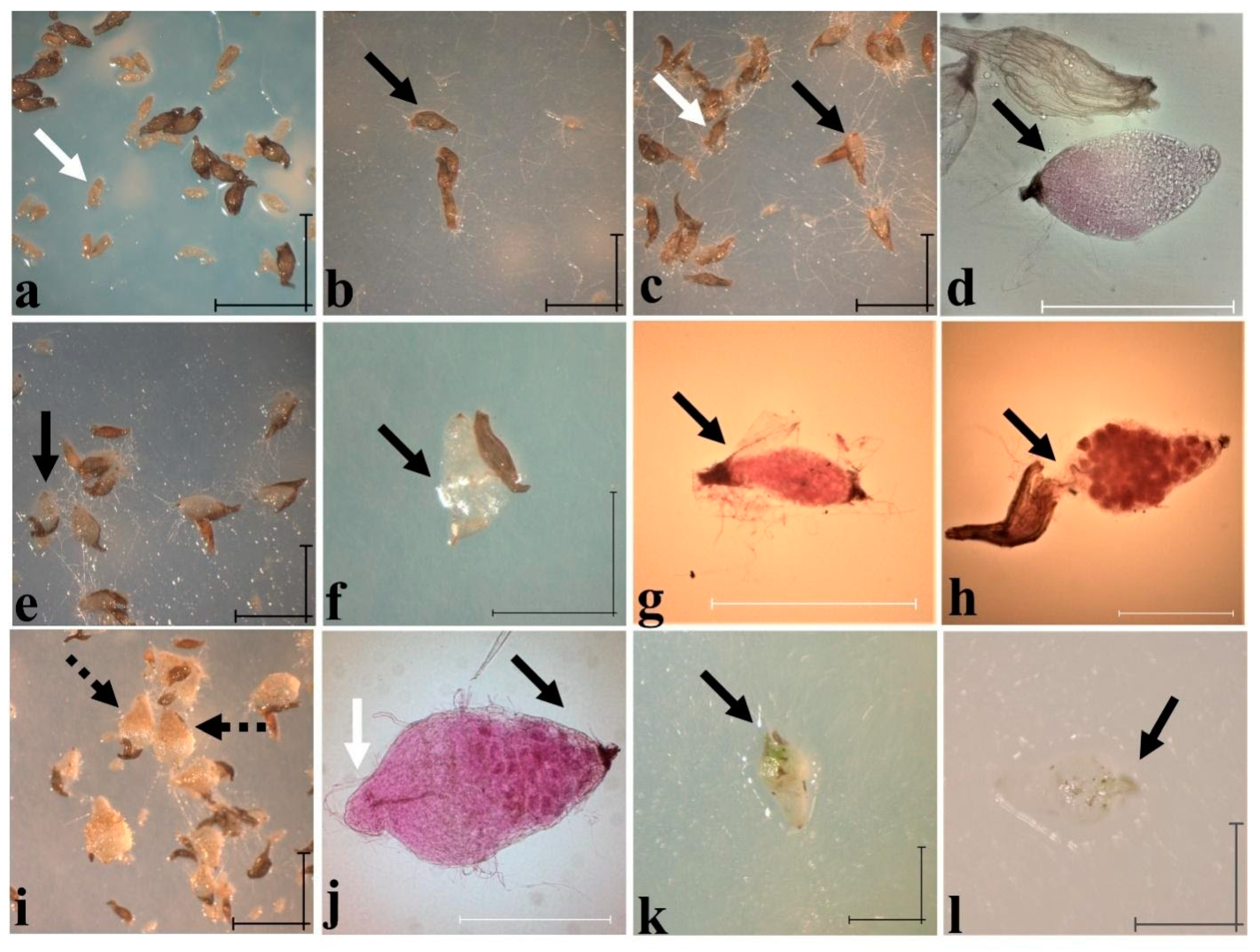
Symbiotic germination was tested on C. irapeanum seeds that were obtained from a population located in an oak forest in Puebla State. Two mycorrhizal endophytes (e.g., Epulorhiza spp.) that were isolated from adult plants from two different sites (e.g., a Quercus forest from Mexico State and a TDF from Veracruz State) were tested, and they exhibited 35% and 20% germination rates, respectively (Table 2). While the isolate from Mexico State promoted a higher germination percentage, the Epulorhiza isolate from Veracruz promoted a more compatible and advanced developmental stage (Figure 11). Shimura and Koda [43] state that symbiotic germination begins after some weeks of incubation in C. macranthos var. rebunense after cool storage, at which time the embryo is imbibed and develops rhizoids that serve as the entry points for fungal colonization. In the case of C. irapeanum, rhizoids did not develop under either asymbiotic or symbiotic conditions (Figure 11a–f), with mycorrhizal colonization likely beginning from the micropylar end (Figure 11d). This colonization is also suitable for water and nutrient absorption [75,76], even if suspensor cells are degraded in the C. calceolus and Mesoamerican hybrid Cypripedium × fred-mulleri to form spherical embryos [18,67]. However, neither of the two identified isolates allow protocorms to develop seedlings (stage 6), as in asymbiotic germination (Figure 10). This incompatibility after symbiotic germination is observed when complete cells that form the protocorm are invaded by fungi, which prevents the apical meristematic cells from undergoing division and growth (Figure 11f–h). This phenomenon could be due to the origin of the isolates, since all were obtained from adult plants [51] or because Cypripedium compatibility with mycorrhizal fungi is low under natural conditions and resulted in the low recruitment observed. Shimura and Koda [43] achieved seedling formation with shoots after cultivation of symbiotic germinated protocorms in an antifungal medium, which indicated that autotrophic plants cannot regulate the symbiotic balance. Other attempts to obtain symbiotic plants have been reported and were without success [65,77].
By comparing asymbiotic and symbiotic development under in vitro conditions, imbibition, rupture of the testa, and tissue differentiation (polarization) occur at similar times after sowing. The multiple lateral shoot induction observed in asymbiotic seedlings produced in vitro could be a consequence of growth under an artificial environment because in situ seedlings developed only one shoot. Unfortunately, there is not yet a successful protocol for propagating lady slipper orchids under either approach.
6. Final Remarks
Mesoamerican Cypripedium species represent an interesting group for study due to their phylogenetic importance as a sister ancestral group to the rest of the genus and because all Cypripedium species are endangered and have some degree of importance. As seen here, their mycorrhizal preferences differ from other species in the genus, and it is probable that they have ecological preferences and, consequently, their morphological variations could be derived from these. The high endangered status of the remaining populations prompts the generation of information on the OMF that promotes in situ germination to develop priority mechanisms for the conservation of hotspot habitats.
Since orchids depend on symbiotic associations, conservation of these species depends on an understanding of the biological and ecological factors that drive the distributions of mainly fungal partners [78,79]. Some aspects may be more determinant than others in species establishment, such as the distributions of suitable mycorrhizal germination-promoting fungi [80]. This contribution provides a partial view of some aspects of the biology of Mesoamerican Cypripedium; however, the successful pollinators, in situ symbiotic germination, and OMF specificity remain unknown. The destruction of habitats urges ex situ conservation strategies that must include not only seed collections but also mycorrhizal fungi that can promote compatible germination or facilitate adaptation. Clarifying the structures of the mycorrhizal fungal communities and isolating germination-promoting fungi would help to determine suitable habitats to conserve or re-establish the studied species and the viability and persistence of in situ seed banks. Asymbiotic propagation in suitable media is a promising tool, but little is known about asymbiotic plant ex vitro adaptation. Both approaches would ensure long-term conservation of these highly endangered species before most of their habitats disappear.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11121554/s1, Supplementary Table S1. Habitat characteristics of Cypripedium spp. remnant populations used for projection of niche ecological model. Supplementary Table S2. Some studies in Cypripedium endophytes using metabarcoding or traditional approaches, entries are ordered in chronological descendent order.
Author Contributions
Conceptualization, M.M.-C. and M.P.O.-L.; formal analysis, M.M.-C.; investigation, M.M.-C. and M.P.O.-L.; writing—original draft preparation, M.M.-C. and M.P.O.-L.; writing—review and editing, M.M.-C. and M.P.O.-L.; project administration and funding acquisition, M.P.O.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the project SEP-CONACYT-ANUIES-ECOS-NORD-FRANCIA 299021, DGAPA-PAPIIT IT-201422 and Posgrado en Ciencias Biológicas from the Universidad Nacional Autónoma de México.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the text and supplemental data. The data presented in this study are available on request from the corresponding author.
Acknowledgments
Authors thank Gerardo Salazar Chávez from Instituto de Biología UNAM for the database of orchid distribution, and Jesus Colín-Rivera, undergraduate student, for in situ baiting germination material. For field assistance, we thank David Moreno Martínez and Verenice Morales from Universidad Veracruzana, Eneida Villavicencio Enríquez from Jardín Botánico Louise Wardle de Camacho, Africam Safari, Quetzal Tonalli Cruz Fernández from UMA Coatzontecoxochitl, and Octavio Gabriel Suarez from UMA Orquideario La Encantada. We also thank Iris Suarez Quijada for laboratory assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chase, M.W. Classification of Orchidaceae in the Age of DNA Data. Curtis’s Bot. Mag. 2005, 22, 2–7. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Luo, Y.B.; Liu, Z.J.; Wang, X.Q. Evolution and biogeography of the slipper orchids: Eocene vicariance of the conduplicate genera in the old and New World tropics. PLoS ONE 2012, 7, e38788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunch, W.D.; Cowden, C.C.; Wurzburger, N.; Shefferson, R.P. Geography and soil chemistry drive the distribution of fungal associations in lady’s slipper orchid, Cypripedium acaule. Botany 2013, 91, 850–856. [Google Scholar] [CrossRef]
- Cribb, P.J. The Genus Cypripedium, 1st ed.; Green, P., Ed.; Timber Press: Portland, OR, USA, 1997; ISBN 9780881924039. [Google Scholar]
- Gargiulo, R.; Pironon, S.; Zheleznaya, E.; Sanchez, M.D.; Balázs, Z.R.; Podar, D.; Wilkinson, T.; Jäkäläniemi, A.; Kull, T.; Väre, H.; et al. Phylogeography and post-glacial dynamics in the clonal-sexual orchid Cypripedium calceolus L. J. Biogeogr. 2019, 46, 526–538. [Google Scholar] [CrossRef]
- Kellof, C.L.; Kass, L.B. Databasing and Georeferencing Historical Collections to Discover Potential Sites for Rare and Endangered Plants of New York, U.S.A. J. Bot. Res. Inst. Tex. J. Bot. Res. 2018, 12, 323–368. [Google Scholar] [CrossRef]
- Li, J.; Liu, Z.; Salazar, G.A.; Bernhardt, P.; Perner, H.; Tomohisa, Y.; Jin, X.; Chung, S.; Luo, Y. Molecular phylogeny of Cypripedium (Orchidaceae: Cypripedioideae) inferred from multiple nuclear and chloroplast regions. Mol. Phylogenet. Evol. 2011, 61, 308–320. [Google Scholar] [CrossRef]
- Pérez-García, E.; Mó, E. Die Cypripedioideae von Mesoamerika. Teil 1a Cypripedium. Die Orchidee. 2014, 65, 476–482. [Google Scholar]
- Unruh, S.A.; McKain, M.R.; Lee, Y.I.; Yukawa, T.; McCormick, M.K.; Shefferson, R.P.; Smithson, A.; Leebens-Mack, J.H.; Pires, J.C. Phylotranscriptomic Analysis and Genome Evolution of the Cypripedioideae (Orchidaceae). Am. J. Bot. 2018, 105, 631–640. [Google Scholar] [CrossRef] [Green Version]
- CITES Appendices | CITES. Available online: https://cites.org/eng/app/appendices.php (accessed on 19 August 2020).
- Mcgough, H.N.; Roberts, D.L.; Brodie, C. CITES y Las Zapatillas de Venus. Royal Botanic Gardens, Kew, United Kingdom. 2006. Available online: https://www.yumpu.com/es/document/read/14398272/cites-y-las-zapatillas-de-venus-royal-botanic-gardens-kew (accessed on 19 August 2020).
- Hagsater, E.; Soto Arenas, M.A.; Salazar, G.; Machorro, J.; López Rosas, M.; Dressler, R. Orchids of Mexico, 1st ed.; Productos Farmacéuticos, S.A. de C.V.; Instituto Chinoin & Redacta: Ciudad de México, México, 2005; ISBN 968788908X. [Google Scholar]
- Soto-Arenas, M.A.; Solano-Gómez, R.A. Ficha Técnica de Cypripedium dickinsonianum. In Información Actualizada Sobre las Especies de Orquídeas del PROY-NOM-059-ECOL-2000; Soto-Arenas, M.A., Solano Gómez, R., Eds.; SNIB-CONABIO: Ciudad de México, Mexico, 2007; pp. 1–7. [Google Scholar]
- Soto-Arenas, M.A.; Solano-Gómez, R.A. Ficha Técnica de Cypripedium irapeanum. In Información Actualizada Sobre las Especies de Orquídeas del PROY-NOM-059-ECOL-2000; Soto-Arenas, M.A., Solano Gómez, R., Eds.; SNIB-CONABIO: Ciudad de México, Mexico, 2007; pp. 1–12. [Google Scholar]
- Soto Arenas, M.Á.; Gómez, R.S.; Hágsater, E. Risk of Extinction and Patterns of Diversity Loss in Mexican Orchids. Lankesteriana 2007, 7, 114–121. [Google Scholar] [CrossRef]
- González-Tamayo, J.R.; Hernández, L. Las Orquídeas del Occidente de México; González-Tamayo, J.R., Hernández, H.L., Eds.; COECYTJAL: Guadalajara, México, 2010; Volume 1, ISBN 9786070040412. [Google Scholar]
- McVaugh, R. Flora Novo-Galiciana: A Descriptive Account of the Vascular Plants of Western Mexico: Orchidaceae, 1st ed.; Anderson, W.R., Ed.; University of Michigan Press: Ann Arbor, MI, USA, 1985; Volume 16, ISBN 0472048163. [Google Scholar]
- Szlachetko, D.L.; Kolanowska, M.; Muller, F.; Vannini, J.; Rojek, J.; Górniak, M. First Guatemalan Record of Natural Hybridisation between Neotropical Species of the Lady’s Slipper Orchid (Orchidaceae, Cypripedioideae). PeerJ 2017, 5, e4162. [Google Scholar] [CrossRef]
- Pérez-García, E.; Mó-Mó, E. Die Cypripedioideae von Mesoamerika. Teil 2b Phragmipedium. Die Orchidee. 2015, 66, 216–225. [Google Scholar]
- Rankou, H.; Salazar, G.A. Conservation Status of Cypripedium irapeanum. Available online: https://www.iucnredlist.org/species/43316630/43327669 (accessed on 19 August 2020).
- NOM-059-SEMARNAT-2010, N.O.M.; NORMA Oficial Mexicana NOM-059-SEMARNAT-2010. Diario Oficial de la Federación: México, 2019. Available online: https://www.dof.gob.mx/normasOficiales/4254/semarnat/semarnat.htm (accessed on 1 April 2022).
- California Academy of Sciences, 2008. Una Comunidad Para Naturalistas. INaturalist. Available online: https://www.inaturalist.org/ (accessed on 12 March 2020).
- INEGI Instituto Nacional de Estadística y Geografía. Available online: http://www.inegi.org.mx/ (accessed on 4 November 2018).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Proal, J.P. Las Secuelas del Boom Mezcalero: Sabor Adulterado, Daños al Ecosistema, Corrupción. Available online: https://www.proceso.com.mx/428051/las-secuelas-del-boom-mezcalero-sabor-adulterado-danos-al-ecosistema-corrupcion?fbclid=IwAR3ENT16jaT081WKYWFcgjQ0EsSRCUH6xgvvp116vxgr2_HrErewDMFzrGk (accessed on 26 May 2022).
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Available online: http://worldclim.org/version2 (accessed on 26 May 2022).
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions; (open-source software) (Version 3.4.1); Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 14 August 2018).
- Global Soil Data Task Group Global Gridded Surfaces of Selected Soil Characteristics (IGBP-DIS). Available online: https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=569 (accessed on 26 May 2022).
- ESRI. Esri: ArcGIS© Pro (Version 2.5), 2016. Available online: https://support.esri.com/en/products/desktop/arcgis-desktop/arcgis-pro/2-5-1 (accessed on 1 April 2022).
- Liu, H.; Jacquemyn, H.; He, X.; Chen, W.; Huang, Y.; Yu, S.; Lu, Y.; Zhang, Y. The Impact of Human Pressure and Climate Change on the Habitat Availability and Protection of Cypripedium (Orchidaceae) in Northeast China. Plants 2021, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.T. Germination and Seedling Development in Five Species of Cypripedium L. Am. J. Bot. 1943, 30, 199–206. [Google Scholar] [CrossRef]
- Rasmussen, H.N. Terrestrial Orchids, 1st ed.; Danish Institute of Plant and Soil Science, Ed.; Cambridge University Press: New York, NY, USA, 1995; ISBN 9780521451659. [Google Scholar]
- Weldy, T.W.; Mlodozeniec, H.T.; Wallace, L.E.; Case, M.A. The Current Status of Cypripedium kentuckiense (Orchidaceae) Including a Morphological Analysis of a Newly Discovered Population in Eastern Virginia. SIDA Contrib. Bot. 1996, 17, 423–435. [Google Scholar]
- Zhongjian, L.; Lijun, C.; Wenhui, R.; Liqiang, L.; Yuting, Z. Correlation between Numerical Dynamics and Reproductive Behavior in Cypripedium lentiginosum. Acta Ecol. Sin. 2008, 28, 111–121. [Google Scholar] [CrossRef]
- Moreno Camarena, M. Caracterización Molecular de Endofitos Micorrízicos y Germinación Simbiótica de Cypripedium irapeanum (Orchidaceae); Universidad Nacional Autónoma de México: Ciudad de México, Mexico, 2016. [Google Scholar]
- Shefferson, R.P.; Weiß, M.; Kull, T.; Taylor, D.L. High Specificity Generally Characterizes Mycorrhizal Association in Rare Lady’s Slipper Orchids, Genus Cypripedium. Mol. Ecol. 2005, 14, 613–626. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Taylor, D.L.; Weiß, M.; Garnica, S.; McCormick, M.K.; Adams, S.; Gray, H.M.; McFarland, J.W.; Kull, T.; Tali, K.; et al. The Evolutionary History of Mycorrhizal Specificity among Lady’s Slipper Orchids. Evolution 2007, 61, 1380–1390. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Kull, T.; Tali, K.; Kellett, K.M. Linking Vegetative Dormancy to Fitness in Two Long-Lived Herbaceous Perennials. Ecosphere 2012, 3, art13. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Kull, T.; Hutchings, M.J.; Selosse, M.A.; Jacquemyn, H.; Kellett, K.M.; Menges, E.S.; Primack, R.B.; Tuomi, J.; Alahuhta, K.; et al. Drivers of Vegetative Dormancy across Herbaceous Perennial Plant Species. Ecol. Lett. 2018, 21, 724–733. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and Seedling Establishment in Orchids: A Complex of Requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Apolinar, M.; Gutierrez-Paredes, C.C.; Sanchez-Gallen, I.; Aguirre, E.; Perez García, E.A. Ecological Aspects of Cypripedium irapeanum La Llave & Lex., an Endangered Mexican orchid Species. Slipper Orchid Alliance Newsl. 2012, 13, 1–5. [Google Scholar]
- Rasmussen, H.N.; Pedersen, H.Æ. Cypripedium calceolus Germination in situ: Seed Longevity, and Dormancy Breakage by Long Incubation and Cold Winters. Eur. J. Enviromental Sci. 2012, 1, 69–70. [Google Scholar] [CrossRef] [Green Version]
- Shimura, H.; Koda, Y. Enhanced Symbiotic Seed Germination of Cypripedium macranthos Var. rebunense Following Inoculation after Cold Treatment. Physiol. Plant. 2005, 123, 281–287. [Google Scholar] [CrossRef]
- Yuan, L.; Yang, Z.L.; Li, S.Y.; Hu, H.; Huang, J.L. Mycorrhizal Specificity, Preference, and Plasticity of Six Slipper Orchids from South Western China. Mycorrhiza 2010, 20, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Dahlberg, C.J.; Carlgren, K.H.I.; Appelqvist, T. Pollination of the Lady’s Slipper Orchid (Cypripedium calceolus) in Scandinavia—Taxonomic and Conservational Aspects. Nord. J. Bot. 2009, 27, 266–273. [Google Scholar] [CrossRef]
- Sugiura, N.; Fujie, T.; Inoue, K.; Kitamura, K. Flowering Phenology, Pollination, and Fruit Set of Cypripedium macranthos var. rebunense, a Threatened Lady’s Slipper (Orchidaceae). J. Plant Res. 2001, 114, 171–178. [Google Scholar] [CrossRef]
- Suetsugu, K.; Fukushima, S. Pollination Biology of the Endangered Orchid Cypripedium japonicum in a Fragmented Forest of Japan. Plant Species Biol. 2014, 29, 294–299. [Google Scholar] [CrossRef]
- Shefferson, R.P.; Bunch, W.; Cowden, C.C.; Lee, Y.I.; Kartzinel, T.R.; Yukawa, T.; Downing, J.; Jiang, H. Does Evolutionary History Determine Specificity in Broad Ecological Interactions? J. Ecol. 2019, 107, 1582–1593. [Google Scholar] [CrossRef]
- Colin Rivera, J.; Ortega-Larrocea, M.P. Colonización Micorrizica de Cypripedium irapeanum La Llave & Lex. (Orchidaceae) En Un Bosque de Quercus. In Conservación de orquídeas en México; Téllez-Velasco, M.d.l.A., Ed.; Instituto de Biología, UNAM: Ciudad de México, México, 2012; pp. 186–192. [Google Scholar]
- Porras-Alfaro, A.; Bayman, P. Mycorrhizal Fungi of Vanilla: Diversity, Specificity and Effects on Seed Germination and Plant Growth. Mycologia 2007, 99, 510–525. [Google Scholar] [CrossRef]
- Shimura, H.; Sadamoto, M.; Matsuura, M.; Kawahara, T.; Naito, S.; Koda, Y. Characterization of Mycorrhizal Fungi Isolated from the Threatened Cypripedium macranthos in a Northern Island of Japan: Two Phylogenetically Distinct Fungi Associated with the Orchid. Mycorrhiza 2009, 19, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Clements, M.A.; Muir, H.; Cribb, P.J. A Preliminary Report on the Symbiotic Germination of European Terrestrial Orchids. Kew Bull. 1986, 41, 437. [Google Scholar] [CrossRef]
- Taylor, D.L.; McCormick, M.K. Internal Transcribed Spacer Primers and Sequences for Improved Characterization of Basidiomycetous Orchid Mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Egidi, E.; May, T.W.; Franks, A.E. Seeking the Needle in the Haystack: Undetectability of Mycorrhizal Fungi Outside of the Plant Rhizosphere Associated with an Endangered Australian Orchid. Fungal Ecol. 2018, 33, 13–23. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Y.C.; Qin, L.Y.; He, H.Y.; Yu, X.L.; Yang, M.Z.; Zhang, H.B. Dynamics of Fungal Communities during Gastrodia elata Growth. BMC Microbiol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Oja, J.; Kohout, P.; Tedersoo, L.; Kull, T.; Kõljalg, U. Temporal Patterns of Orchid Mycorrhizal Fungi in Meadows and Forests as Revealed by 454 Pyrosequencing. New Phytol. 2015, 205, 1608–1618. [Google Scholar] [CrossRef]
- Valdés, M.; Bautista Guerrero, H.; Martínez, L.; Víquez, R. The Root Colonizing Fungi of the Terrestrial Orchid Cypripedium irapeanum. Lankesteriana 2011, 11, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Zelmer, C.D.; Cuthbertson, L.; Currah, R.S. Fungi Associated with Terrestrial Orchid Mycorrhizas, Seeds and Protocorms. Mycoscience 1996. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T. ITS Primers with Enhanced Specifity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.; Yokoya, K.; Kendon, J.P.; Sarasan, V. Diversity of Root-Associated Culturable Fungi of Cephalanthera rubra (Orchidaceae) in Relation to Soil Characteristics. PeerJ 2020, 2020, e8695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, M.M.; Stewart, J. Re-Establishment of the Ladt’s Slipper Orchid (Cypripedium calceolus L.) in Britain. Bot. J. Linn. Soc. 1998, 126, 173–181. [Google Scholar]
- Barsberg, S.; Rasmussen, H.N.; Kodah, N. Composition of Cypripedium calceolus (Orchidaceae) Seeds Analyzed by Attenuated Total Reflectance Ir Spectroscopy: In Search of Understanding Longevity in the Ground. Am. J. Bot. 2013, 100, 2066–2073. [Google Scholar] [CrossRef]
- Zeng, S.; Zhang, Y.; Teixeira da Silva, J.A.; Wu, K.; Zhang, J.; Duan, J. Seed Biology and in vitro Seed Germination of Cypripedium. Crit. Rev. Biotechnol. 2014, 34, 358–371. [Google Scholar] [CrossRef]
- Lee, Y.I.; Chung, M.C.; Yeung, E.C.; Lee, N. Dynamic Distribution and the Role of Abscisic Acid during Seed Development of a Lady’s Slipper Orchid, Cypripedium formosanum. Ann. Bot. 2015, 116, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lee, Y.I.; Deng, L.; Zhao, S. Asymbiotic Germination of Immature Seeds and the Seedling Development of Cypripedium macranthos Sw., an Endangered Lady’s Slipper Orchid. Sci. Hortic. 2013, 164, 130–136. [Google Scholar] [CrossRef]
- Pedersen, H.A.; Rasmussen, H.N.; Kahandawala, I.M.; Fay, M.F. Genetic Diversity, Compatibility Patterns and Seed Quality in Isolated Populations of Cypripedium calceolus (Orchidaceae). Conserv. Genet. 2012, 13, 89–98. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, M.C.; Lee, Y.I. In vitro Germination and Low-Temperature Seed Storage of Cypripedium lentiginosum P.J. Cribb & S.C. Chen, a Rare and Endangered Lady’s Slipper Orchid. Sci. Hortic. 2017, 225, 471–479. [Google Scholar] [CrossRef]
- SIGMA Phytamax TM. Orchid Maintenance Medium. Available online: https://www.sigmaaldrich.com/catalog/product/sigma/p6668?lang=es®ion=MX (accessed on 26 May 2022).
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Norstog, K. Induction of Embryolike Structures by Kinetin in Cultured Barley Embryos. Dev. Biol. 1970, 23, 665–670. [Google Scholar] [CrossRef]
- Hsu, R.C.C.; Lee, Y.I. Seed Development of Cypripedium debile Rchb. f. in Relation to Asymbiotic Germination. HortScience 2012, 47, 1495–1498. [Google Scholar] [CrossRef] [Green Version]
- Yeung, E.C. A Perspective on Orchid Seed and Protocorm Development Background. Bot. Stud. 2017, 58. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.F. Orchid Conservation: How Can We Meet the Challenges in the Twenty-First Century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [Green Version]
- Jakubska-Busse, A.; Tsiftsis, S.; Śliwiński, M.; Křenová, Z.; Djordjević, V.; Steiu, C.; Kolanowska, M.; Efimov, P.; Hennigs, S.; Lustyk, P.; et al. How to Protect Natural Habitats of Rare Terrestrial Orchids Effectively: A Comparative Case Study of Cypripedium calceolus in Different Geographical Regions of Europe. Plants 2021, 10, 404. [Google Scholar] [CrossRef]
- Swarts, N.D.; Batty, A.L.; Hopper, S.; Dixon, K.W. Does Integrated Conservation of Terrestrial Orchids Work? Lankesteriana 2007, 7, 219–222. [Google Scholar] [CrossRef] [Green Version]
- McCormick, M.K.; Lee Taylor, D.; Juhaszova, K.; Burnett, R.K.; Whigham, D.F.; O’Neill, J.P. Limitations on Orchid Recruitment: Not a Simple Picture. Mol. Ecol. 2012, 21, 1511–1523. [Google Scholar] [CrossRef]
Figure 1.
Mesoamerican Cypripedium: (a) C. dickinsonianum, (b) C. irapeanum and (c) C. molle, pictures taken by Javier Fortanelli, Mauricio Moreno, and Octavio Gabriel in the years 2014, 2010 and 2019, respectively.
Figure 1.
Mesoamerican Cypripedium: (a) C. dickinsonianum, (b) C. irapeanum and (c) C. molle, pictures taken by Javier Fortanelli, Mauricio Moreno, and Octavio Gabriel in the years 2014, 2010 and 2019, respectively.

Figure 2.
Actual distribution of Mesoamerican Cypripediums from herbarium data. Cypripedium irapeanum = yellow points, C. molle = red points, and C. dickinsonianum = blue points.
Figure 2.
Actual distribution of Mesoamerican Cypripediums from herbarium data. Cypripedium irapeanum = yellow points, C. molle = red points, and C. dickinsonianum = blue points.

Figure 3.
Destroyed habitats of Mesoamerican Cypripedium molle in Oaxaca State (a–d) and C. irapeanum in Nayarit State (e–f). (a) Land use change for Agave angustifolia cultivation for mezcal production (white arrow indicates a remaining C. molle plant). (b) Deforestation of a pine forest in the transition from oak to cloud forest habitat. (c) Fire destruction of oak forest habitat in limestone. (d) Habitat destruction by an outdoor dump. (e) Population vulnerability next to the road subject to any perturbation process. (f) Land use change of deciduous forest transitions for mango cultivars (white arrow shows the disappeared population). (a–e), photos taken by Mauricio Moreno in 2019; (f), INEGI [23] and Google Earth ©, 2020.
Figure 3.
Destroyed habitats of Mesoamerican Cypripedium molle in Oaxaca State (a–d) and C. irapeanum in Nayarit State (e–f). (a) Land use change for Agave angustifolia cultivation for mezcal production (white arrow indicates a remaining C. molle plant). (b) Deforestation of a pine forest in the transition from oak to cloud forest habitat. (c) Fire destruction of oak forest habitat in limestone. (d) Habitat destruction by an outdoor dump. (e) Population vulnerability next to the road subject to any perturbation process. (f) Land use change of deciduous forest transitions for mango cultivars (white arrow shows the disappeared population). (a–e), photos taken by Mauricio Moreno in 2019; (f), INEGI [23] and Google Earth ©, 2020.

Figure 4.
Distribution of suitable habitats of Mesoamerican Cypripedium based on the most reliable Maxent model. Rasters generated in Maxent [27]. Maps were generated in ArcGIS© 10.5 (ESRI, https://www.esri.com/en-us/home (accessed on 1 April 2022)) (a) C. dickinsonianum, (b) C. irapeanum, and (c) C. molle.
Figure 4.
Distribution of suitable habitats of Mesoamerican Cypripedium based on the most reliable Maxent model. Rasters generated in Maxent [27]. Maps were generated in ArcGIS© 10.5 (ESRI, https://www.esri.com/en-us/home (accessed on 1 April 2022)) (a) C. dickinsonianum, (b) C. irapeanum, and (c) C. molle.

Figure 5.
In situ development of Cypripedium molle (a,b) in an oak forest in Oaxaca State and C. irapeanum (c–f) in an oak forest in Mexico State and in a tropical dry forest in Veracruz State. (a) Long, little-branched roots longer than 30 cm under abundant litter (white arrow). (b) Achlorophyllous shoot (red arrow) of an emerging plant sprouting due to the deep litter where roots develop in the humic horizon. (c) Lateral shoot emerging from a near meristematic node (green arrow) and short rhizome development with roots surrounded by mycelial cords (dashed red arrow) with short-distance exploration mycelium (double-line red arrow). (d) Multiple points of emergence of annual plants showing a node with growth of several years (white arrows) and the remainder of a dried shoot from the previous year (blue arrow). (e) Plants usually associated with Bletia orchids (B. purpurea, white arrow). (f) Association with an eight-year-old Bletia punctata plant showing the emerging shoots of the first two submerged bulbs. These belong to the first and second bulbs that developed after germination and the emerging shoots after eight years (white arrow). A dried C. irapeanum shoot from the previous year next to a new emerging shoot (yellow arrow). Photos taken by Mauricio Moreno in the year 2019.
Figure 5.
In situ development of Cypripedium molle (a,b) in an oak forest in Oaxaca State and C. irapeanum (c–f) in an oak forest in Mexico State and in a tropical dry forest in Veracruz State. (a) Long, little-branched roots longer than 30 cm under abundant litter (white arrow). (b) Achlorophyllous shoot (red arrow) of an emerging plant sprouting due to the deep litter where roots develop in the humic horizon. (c) Lateral shoot emerging from a near meristematic node (green arrow) and short rhizome development with roots surrounded by mycelial cords (dashed red arrow) with short-distance exploration mycelium (double-line red arrow). (d) Multiple points of emergence of annual plants showing a node with growth of several years (white arrows) and the remainder of a dried shoot from the previous year (blue arrow). (e) Plants usually associated with Bletia orchids (B. purpurea, white arrow). (f) Association with an eight-year-old Bletia punctata plant showing the emerging shoots of the first two submerged bulbs. These belong to the first and second bulbs that developed after germination and the emerging shoots after eight years (white arrow). A dried C. irapeanum shoot from the previous year next to a new emerging shoot (yellow arrow). Photos taken by Mauricio Moreno in the year 2019.

Figure 6.
In situ germination of Cypripedium irapeanum from Veracruz State in a tropical dry forest (a,b) and an oak forest in Mexico State (c–h). (a) Seedling less than 3 cm long. (b). Emergence of multiple large roots from seedlings (white arrow). (c) Seed imbibition (white arrow) after the second month of baiting placement (upper right picture), showing that the sand grain sizes are larger than those of the seeds and brown fungal hyphae (scale bar = 500 µm). (d,e) Two-month achlorophyllous protocorms in the baiting showing development of the shoot apical apex (scale bar = 500 µm). (f) Germinated seed after 10 years of baiting in August 2018 with a root that is several times larger than the undeveloped shoot (white arrow) (scale bar = 16 mm). (g) Detail of a root emerging from an undeveloped shoot of the same seedling showing testa remains (white arrow) (scale bar = 500 µm). (h) Comparative size of seedlings regarding baiting dispositive (see (c)) (scale bar = 2 mm). Photos taken by Mauricio Moreno (a,b) in 2020; Jesus Colín (c) in 2012 and M.P. Ortega (d–h) in 2018.
Figure 6.
In situ germination of Cypripedium irapeanum from Veracruz State in a tropical dry forest (a,b) and an oak forest in Mexico State (c–h). (a) Seedling less than 3 cm long. (b). Emergence of multiple large roots from seedlings (white arrow). (c) Seed imbibition (white arrow) after the second month of baiting placement (upper right picture), showing that the sand grain sizes are larger than those of the seeds and brown fungal hyphae (scale bar = 500 µm). (d,e) Two-month achlorophyllous protocorms in the baiting showing development of the shoot apical apex (scale bar = 500 µm). (f) Germinated seed after 10 years of baiting in August 2018 with a root that is several times larger than the undeveloped shoot (white arrow) (scale bar = 16 mm). (g) Detail of a root emerging from an undeveloped shoot of the same seedling showing testa remains (white arrow) (scale bar = 500 µm). (h) Comparative size of seedlings regarding baiting dispositive (see (c)) (scale bar = 2 mm). Photos taken by Mauricio Moreno (a,b) in 2020; Jesus Colín (c) in 2012 and M.P. Ortega (d–h) in 2018.

Figure 7.
Mycorrhizal root colonization of Mesoamerican Cypripedium. View of one of the main roots (black arrows) and secondary roots (white arrows) of Cypripedium irapeanum (a–e) and C. molle (f–j). Cortical colonization by mycorrhizal fungi occurs in patches on the main roots with long sections without fungal colonization and starch grains (b,c) and some with very dense colonization with digested pelotons (f,g). SR are always densely colonized with undigested and partially digested pelotons (d,e and h,i). Bars in c, e, g, i = 10 µm and bars in b, d, f, h = 100 µm. Bar in a = 10 cm. Photos taken by Mauricio Moreno in year 2019.
Figure 7.
Mycorrhizal root colonization of Mesoamerican Cypripedium. View of one of the main roots (black arrows) and secondary roots (white arrows) of Cypripedium irapeanum (a–e) and C. molle (f–j). Cortical colonization by mycorrhizal fungi occurs in patches on the main roots with long sections without fungal colonization and starch grains (b,c) and some with very dense colonization with digested pelotons (f,g). SR are always densely colonized with undigested and partially digested pelotons (d,e and h,i). Bars in c, e, g, i = 10 µm and bars in b, d, f, h = 100 µm. Bar in a = 10 cm. Photos taken by Mauricio Moreno in year 2019.

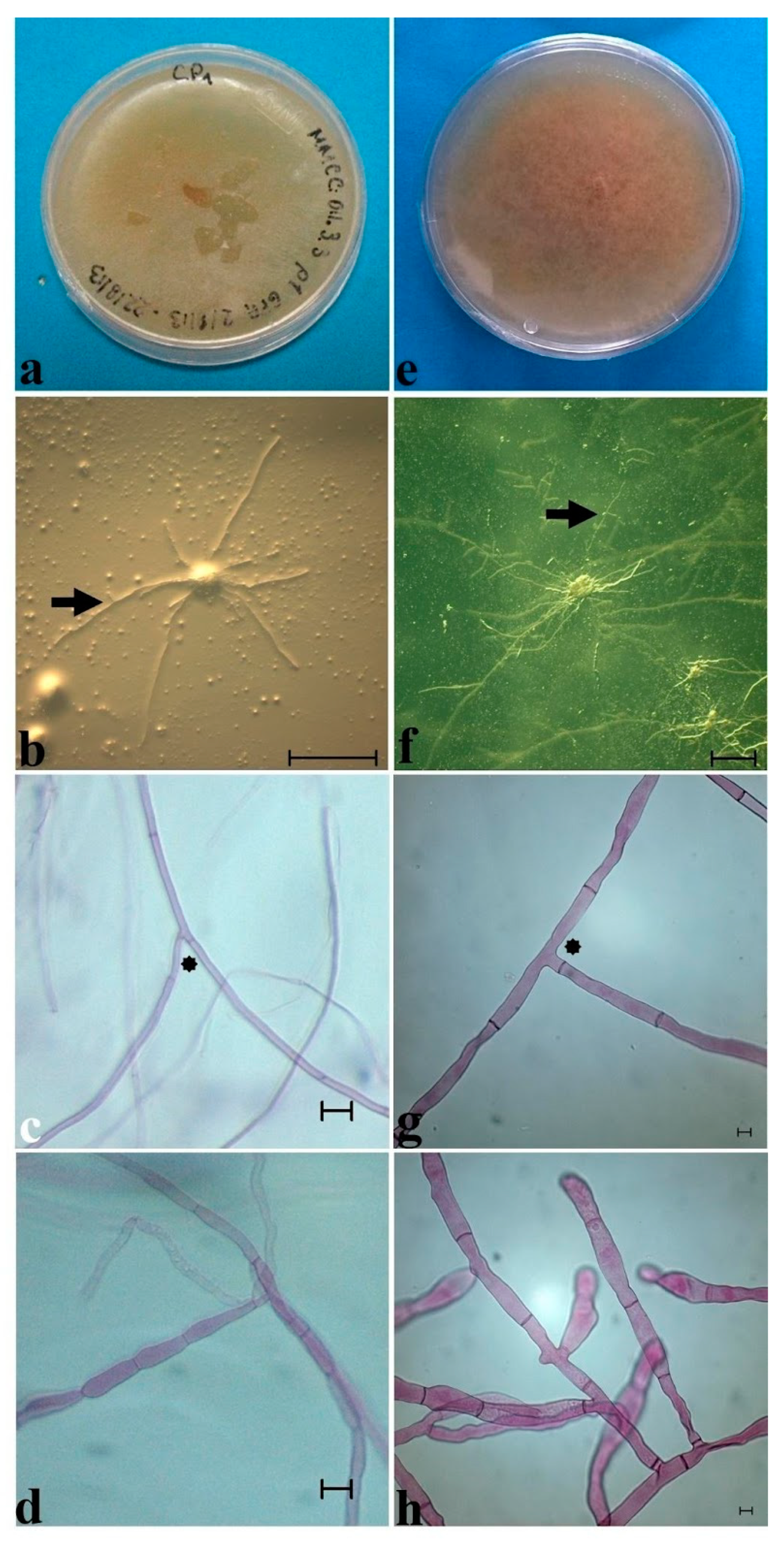
Figure 8.
Mycorrhizal isolates of Mesoamerican Cypripedium. Isolate from Cypripedium irapeanum (a–d) and from C. molle (e–h). (a) Colony view of Epulorhiza sp. (Tulasnella sp.) from a tropical dry forest in Veracruz State showing a submerged mycelium, yellow-creamy color, and waxy surface. (b) Superficial growth after 5 days of incubation of straight hyphae obtained from a peloton after sowing on GPA medium showing 90° bifurcation of the hyphae (black arrow). Bar = 100 µm. (c) Mycelium from an isolated culture showing less than 4 µm hyphae and straight basal septa (black asterisk). Bar = 10 µm. (d) Monilioid cells. Bar = 10 µm. (e) Colony view of Ceratorhiza sp. (Ceratobasidium sp.) that was isolated from a Quercus forest in Oaxaca State showing an aerial mycelium, brownish-creamy color, and cottony superficial aspect. (f) View of hyphae growing from a submerged peloton with a knobby appearance after 3 days of incubation. Bar = 10 µm. (g) Mycelium from an isolated culture showing hyphae longer than 4 µm and constrained basal septa (black asterisk). Bar = 10 µm. (h) Monilioid cells. Bar = 10 µm. Photos taken by Mauricio Moreno in 2020.
Figure 8.
Mycorrhizal isolates of Mesoamerican Cypripedium. Isolate from Cypripedium irapeanum (a–d) and from C. molle (e–h). (a) Colony view of Epulorhiza sp. (Tulasnella sp.) from a tropical dry forest in Veracruz State showing a submerged mycelium, yellow-creamy color, and waxy surface. (b) Superficial growth after 5 days of incubation of straight hyphae obtained from a peloton after sowing on GPA medium showing 90° bifurcation of the hyphae (black arrow). Bar = 100 µm. (c) Mycelium from an isolated culture showing less than 4 µm hyphae and straight basal septa (black asterisk). Bar = 10 µm. (d) Monilioid cells. Bar = 10 µm. (e) Colony view of Ceratorhiza sp. (Ceratobasidium sp.) that was isolated from a Quercus forest in Oaxaca State showing an aerial mycelium, brownish-creamy color, and cottony superficial aspect. (f) View of hyphae growing from a submerged peloton with a knobby appearance after 3 days of incubation. Bar = 10 µm. (g) Mycelium from an isolated culture showing hyphae longer than 4 µm and constrained basal septa (black asterisk). Bar = 10 µm. (h) Monilioid cells. Bar = 10 µm. Photos taken by Mauricio Moreno in 2020.

Figure 9.
Phylogenetic relationships of endophytes isolated from Mesoamerican Cypripedium. The pink clade represents isolates from Cypripedium irapeanum from a tropical dry forest and Quercus forest. The blue clade contains the isolate from C. molle, which was inferred by using the maximum likelihood method and the Tamura-Nei model log likelihood of –3241.26. The branch length labels represent the bootstrap proportions. Analyses were conducted in Geneious (2021.0.3).
Figure 9.
Phylogenetic relationships of endophytes isolated from Mesoamerican Cypripedium. The pink clade represents isolates from Cypripedium irapeanum from a tropical dry forest and Quercus forest. The blue clade contains the isolate from C. molle, which was inferred by using the maximum likelihood method and the Tamura-Nei model log likelihood of –3241.26. The branch length labels represent the bootstrap proportions. Analyses were conducted in Geneious (2021.0.3).

Figure 10.
Asymbiotic in vitro germination of Cypripedium irapeanum in Norstog medium. (a) Development stage 0 on day of sowing (black arrows show immature and undeveloped seeds and white arrows show mature seeds). (b) Imbibed seeds two days after sowing (das) (white arrow) (development stage 1). (c) Imbibed seed with rupture of testa 12 das (white arrow) (development stage 2). (d) Protocorm polarization with apical meristem at 45 das (upper structure) (developmental stage 3). (e) Protocorm development with leaf blade at 54 das (developmental stage 4). (f) Pre-seedling showing radical meristem (black dashed arrow) and a long achlorophyllous leaf 96 das (developmental stage 5). Other chlorophyll pre-seedlings with two leaf blades are located at the bottom left. (g) Seedling development with two incipient roots at 122 das (black dashed arrow) (developmental stage 6). (h) Seedling 187 das with four leaves and a greater ratio between root and shoot. (i) Seedling 300 das with lateral shoots and remnant testa (black dashed arrow) from the first shoot and two radical sprouts. The white arrow shows two oxidized pre-seedlings at stage 4. (j) Four seedlings 300 das with long roots and one seedling with two shoots (black dashed arrow). The white arrow shows a protocorm with irregular protuberances, such as rhizoids, or a protocorm-like body (PLB). (k) Seedling at 365 das with three shoots and two elongated and exfoliant rhizodermic roots. Scale bars = 100 µm.
Figure 10.
Asymbiotic in vitro germination of Cypripedium irapeanum in Norstog medium. (a) Development stage 0 on day of sowing (black arrows show immature and undeveloped seeds and white arrows show mature seeds). (b) Imbibed seeds two days after sowing (das) (white arrow) (development stage 1). (c) Imbibed seed with rupture of testa 12 das (white arrow) (development stage 2). (d) Protocorm polarization with apical meristem at 45 das (upper structure) (developmental stage 3). (e) Protocorm development with leaf blade at 54 das (developmental stage 4). (f) Pre-seedling showing radical meristem (black dashed arrow) and a long achlorophyllous leaf 96 das (developmental stage 5). Other chlorophyll pre-seedlings with two leaf blades are located at the bottom left. (g) Seedling development with two incipient roots at 122 das (black dashed arrow) (developmental stage 6). (h) Seedling 187 das with four leaves and a greater ratio between root and shoot. (i) Seedling 300 das with lateral shoots and remnant testa (black dashed arrow) from the first shoot and two radical sprouts. The white arrow shows two oxidized pre-seedlings at stage 4. (j) Four seedlings 300 das with long roots and one seedling with two shoots (black dashed arrow). The white arrow shows a protocorm with irregular protuberances, such as rhizoids, or a protocorm-like body (PLB). (k) Seedling at 365 das with three shoots and two elongated and exfoliant rhizodermic roots. Scale bars = 100 µm.

Figure 11.
Symbiotic in vitro germination of Cypripedium irapeanum with two Epulorhiza spp. (Tulasnella spp.) isolates obtained from an oak forest in Mexico State and a tropical deciduous forest in Veracruz State with seeds from Puebla State. (a) Seed imbibition (stage 1) before fungal contact at 2 days after sowing (das) (white arrow shows immature and ungerminated seeds with embryos) with a Mexico State isolate. (b) Mycorrhizal contact during seed imbibition at 12 das with the Veracruz isolate. (c) Symbiotic protocorms surrounded by mycelium (not rhizoids) at stages 2 (white arrow) and 3 developing rhizoids (black arrow) at 12 das with a Veracruz isolate. (d) Protocorm out of testa at 12 das stained with acid fuchsine showing pelotons at the micropillar seed region (black arrow) and hyphae growing in medium. (e,f) Symbiotic protocorms at stages 3 (e) and 4 (f) at 22 and 54 das, respectively, with the Veracruz isolate showing early vitrification and fungal incompatibility (black arrows). (g,h) Histological staining evidence of the fungal incompatibility, where pelotons invade apical meristematic tissue cells (black arrows) at stage 3 (g) and stage 4 (h), both at 109 das with an isolate from Mexico State. (i) Symbiotic protocorms at 187 das showing fungal compatibility that are located in a brownish tissue in the micropylar zone (black dashed arrow) with the Veracruz isolate. (j) Histological view of a stained mycorrhized protocorm with dense pink pelotons (black arrow) at the micropylar pole and with intact leaf vascular bundles (white arrow) at 96 das. (k,l) Symbiotic mixotrophic protocorms at stage 4 (with two foliar sheets) at 72 and 109 das with the Veracruz isolate. Scale bar = 100 µm. Photos taken by Mauricio Moreno in 2015.
Figure 11.
Symbiotic in vitro germination of Cypripedium irapeanum with two Epulorhiza spp. (Tulasnella spp.) isolates obtained from an oak forest in Mexico State and a tropical deciduous forest in Veracruz State with seeds from Puebla State. (a) Seed imbibition (stage 1) before fungal contact at 2 days after sowing (das) (white arrow shows immature and ungerminated seeds with embryos) with a Mexico State isolate. (b) Mycorrhizal contact during seed imbibition at 12 das with the Veracruz isolate. (c) Symbiotic protocorms surrounded by mycelium (not rhizoids) at stages 2 (white arrow) and 3 developing rhizoids (black arrow) at 12 das with a Veracruz isolate. (d) Protocorm out of testa at 12 das stained with acid fuchsine showing pelotons at the micropillar seed region (black arrow) and hyphae growing in medium. (e,f) Symbiotic protocorms at stages 3 (e) and 4 (f) at 22 and 54 das, respectively, with the Veracruz isolate showing early vitrification and fungal incompatibility (black arrows). (g,h) Histological staining evidence of the fungal incompatibility, where pelotons invade apical meristematic tissue cells (black arrows) at stage 3 (g) and stage 4 (h), both at 109 das with an isolate from Mexico State. (i) Symbiotic protocorms at 187 das showing fungal compatibility that are located in a brownish tissue in the micropylar zone (black dashed arrow) with the Veracruz isolate. (j) Histological view of a stained mycorrhized protocorm with dense pink pelotons (black arrow) at the micropylar pole and with intact leaf vascular bundles (white arrow) at 96 das. (k,l) Symbiotic mixotrophic protocorms at stage 4 (with two foliar sheets) at 72 and 109 das with the Veracruz isolate. Scale bar = 100 µm. Photos taken by Mauricio Moreno in 2015.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Micromorphological characteristics of Cypripedium spp. Mycorrhizal endophytes (mean ± standard deviation). Different letters represent significant differences at p ≤ 0.05. See Figure 8.
Table 1.
Micromorphological characteristics of Cypripedium spp. Mycorrhizal endophytes (mean ± standard deviation). Different letters represent significant differences at p ≤ 0.05. See Figure 8.
| Plant Species | Mean Day Growth on PDA Medium | Basal Septa (µm) | Diameter of Hyphae (µm) | Monilioid Cells (µm) | |
|---|---|---|---|---|---|
| Mean ± SD | Width | Length | |||
| Cypripedium irapeanum (Tropical deciduous forest, Veracruz) | 1.14 ± 0.22a | 1.794 ± 1.280a | 3.3 ± 1.1a | 7.7 ± 2.0a | 16.1 ± 4.5a |
| C. irapeanum (Quercus forest, Mexico State) | 0.44 ± 0.25b | 1.439 ± 0.384a | 2.7 ± 0.4a | 7.8 ± 1.2a | 18.5 ± 4.2a |
| C. molle (Quercus forest, Oaxaca) | 2.50 ± 1.20c | 7.368 ± 1.679b | 12.6 ± 1.2b | 20.0 ± 1.9b | 36.2 ± 8.4b |
Table 2.
Asymbiotic (1–4 treatments) and symbiotic (5–6 treatments) germination and development of Cypripedium irapeanum seeds after 256 days of sowing. MS = Murashige and Skoog [73], Modified Phytamax ™ [72], Norstog [59], OMA = Oatmeal agar [52]. Stages can be appreciated in Figure 8 and Figure 9.
Table 2.
Asymbiotic (1–4 treatments) and symbiotic (5–6 treatments) germination and development of Cypripedium irapeanum seeds after 256 days of sowing. MS = Murashige and Skoog [73], Modified Phytamax ™ [72], Norstog [59], OMA = Oatmeal agar [52]. Stages can be appreciated in Figure 8 and Figure 9.
| Treatments | STAGES (%) | |||||
|---|---|---|---|---|---|---|
| E0 | E1 | E2–3 | E4 | E5 | E6 | |
| Immature and Ungerminated Seeds with Embryos | Imbibition | Rupture of the Testa and Polarization of the Embryo | Foliar Elongation | Root Meristematic Differentiation | Seedling | |
| MS | 49 | 99 | 0 | 1 | 0 | 0 |
| Phytamax ™ | 71 | 88 | 10 | 0 | 2 | 0 |
| Norstog | 52 | 91 | 1 | 3 | 1 | 4 |
| OMA | 49 | 86 | 14 | 0 | 0 | 0 |
| TDF isolate | 29 | 81 | 19 | 1 | 0 | 0 |
| Quercus forest isolate | 26 | 65 | 24 | 11 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moreno-Camarena, M.; Ortega-Larrocea, M.P. Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants 2022, 11, 1554. https://doi.org/10.3390/plants11121554
AMA Style
Moreno-Camarena M, Ortega-Larrocea MP. Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species. Plants. 2022; 11(12):1554. https://doi.org/10.3390/plants11121554
Chicago/Turabian StyleMoreno-Camarena, Mauricio, and María Pilar Ortega-Larrocea. 2022. "Mesoamerican Cypripedium: Mycorrhizal Contributions to Promote Their Conservation as Critically Endangered Species" Plants 11, no. 12: 1554. https://doi.org/10.3390/plants11121554
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.