
Ideotype Selection of Perennial Flax (Linum spp.) for Herbaceous Plant Habit Traits

1
Department of Horticultural Science, University of Minnesota, 1970 Folwell Ave, Saint Paul, MN 55108, USA
2
Department of Agronomy & Plant Genetics, University of Minnesota, 1529 Lindig St., Saint Paul, MN 55108, USA
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(12), 3127; https://doi.org/10.3390/agronomy12123127
Submission received: 11 November 2022
/
Revised: 2 December 2022
/
Accepted: 8 December 2022
/
Published: 9 December 2022
(This article belongs to the Special Issue Advances in Plant Genetic Breeding and Molecular Biology)
Abstract
:(1) Background: Annual flax (Linum usitatissimum) and many wild relatives possess showy blue flowers and finely textured foliage. To promote the use of blue-flowered flax as ornamentals, an herbaceous perennial flax breeding program was initiated to develop ideotypes and test their effectiveness. The objectives of this study were to (a) compare traits of interest for herbaceous perennials in breeding populations (CF, oilseed) and accessions of annual/perennial species in a common garden; (b) quantify the impact of selection (direct, indirect) for traits of interest; (c) identify top candidate species for continued breeding using the herbaceous perennial flax crop ideotype; (2) Methods: Recorded traits, based on the perennial flax ideotype, included: flower diameter, flowering period, stem length and diameter, plant width and height, summer and winter survival; (3) Results: OS and CF populations had smaller stem diameters, longer flowering periods, larger plant size, more uniform growth, and improved winter survival compared to wild species. Linum austriacum was the top wild species for the CF breeding ideotype but comparable in performance with L. perenne for the herbaceous perennial flax ideotype; (4) Conclusions: The effect of 1–5 yrs. on selection for target/non-target traits in wild species for future R&D is exemplified with selection and release of an herbaceous/garden ornamental perennial flax for the market.
1. Introduction
The prolific flowers of some perennial flax species make them ideal candidates for ornamental uses, such as garden bedding plants, CF or HP [1,2]. Perennial flax can also fill gaps in the pollinator food supply due to its long flowering period [3]. In a unique collaboration between agronomy and horticulture breeders, the Forever Green Initiative (FGI) perennial flax breeding program is simultaneously pursuing both oilseed, fiber, pollinator, and ornamental breeding objectives (CF, HP or ornamental garden types), allowing for efficiency in breeding and selection of multiple traits simultaneously [3]. These efforts are being facilitated using an ideotype approach; the objectives and goals of the FGI program are previously described [1].
The genus Linum is distributed throughout temperate and subtropical regions of the world (Bolsheva et al., 2017; McDill et al., 2009) and taxonomically classified, based on morphological [4,5,6,7,8,9,10,11,12,13] and molecular traits [4]. The Adenolinum or L. perenne group within sect. Linum [5] is the primary source of perennial flax germplasm being developed by the FGI breeding program. It includes Eurasian species such as L. perenne L. and L. austriacum L. as well as the North American species L. lewisii Pursh [5,10]. Other species such as L. perenne, L. narbonense L., and L. grandiflorum Desf. have ornamental value, although there are few reports on the variation available for ornamental breeding [14,15]. Section Linum also encompasses notable species such as annual flax (L. usitatissimum) and its progenitor L. bienne Mill. (=L. angustifolium Huds.) [5]. Several Mediterranean and southwest Asian species have been cultivated as ornamentals, such as L. hypericifolium Salisb., L. hirsutum L. and L. viscosum L. [5,16,17]. One species within the yellow-flowered clade (sections Linopsis, Syllinum, and Cathartolinum) includes perennial L. flavum L. [5,16,17].
Flax grows a dominant primary shoot (prostrate to erect), with lateral branching occurring primarily from the base to form a crown with an underground taproot. The green, occasionally glabrous, leaves are alternate in phyllotaxy, linear to linear-lanceolate in shape and 15–55 mm in length [17,18]. New flowers open each morning with petal drop starting midday [17,19]. Petals can be shades of blue, white, pink, violet, red, or yellow, depending on the species. Petal veins, stamens, and anthers can all exhibit the same range of color as the petals, with independent inheritance [17]. The fruit of annual flax, produced for oil seed, is a boll or capsule divided into five locules with potential seed set of 2 seeds/locule for a maximum of 10 seeds/boll [20]. There is variation within the genus for all reproductive traits, including sepal and petal shape, flower size and color, seed size and color, and degree of capsule opening [18].
Many species also exhibit sexual dimorphism (heterostyly) and a sporophytic self incompatibility (SI) system, both of which promote outcrossing [21]. In the classic case of distyly, there exist two flower morphs: approach herkogamous (pin), where the styles are longer than the stamens, and reverse herkogamous (thrum) where the styles are shorter than the stamens [22]. In this SI system, only pollen from the reciprocal flower morph will result in successful pollination. There are also species in Linum, such as domesticated annual flax that is self-compatible (SC) and monomorphic homostylous, meaning that styles and stamens are at equal height in all flowers [21]. Finally, there is the unique case of L. lewisii, which is monomorphic approach herkogamous, yet SC [23]. This floral trait is one of the few morphological distinctions between North American L. lewisii and Eurasian L. perenne, which has caused misidentifications [23].
We have previously tested specific traits in perennial flax crop ideotypes for joint horticultural-agronomic breeding objectives [3], cold tolerance and winter hardiness [24], a critical trait for both oilseed (OS) and CF perennial flax, as well as CF postharvest longevity [2]. The traits for the herbaceous perennial (HP) or ornamental garden crop ideotype have yet to be identified and tested, particularly in comparison with CF and OS ideotypes. The objectives of this study were to (1) compare traits of interest for breeding populations (CF, OS, along with HP) and accessions of various annual and perennial species in a common garden; (2) quantify the impact of past selection (direct, indirect) for traits of interest for HP flax ideotypes; (3) identify the top candidate species for continued ornamental flax breeding using identified traits for the HP flax crop ideotype. Recorded traits were based on perennial flax ideotypes and included: flower diameter, flowering period, stem length and diameter, plant width and height, germination by week, summer and winter survival. This trait evaluation tests the assumptions and hypotheses about the perennial flax crop ideotypes previously identified [1]. We hypothesize that the CF and OS selection populations will possess mean trait values exceeding the species accessions for the traits under selection whilst HP trait values may be similar to or differ from CF and OS traits since they have not been previously selected.
2. Materials and Methods
2.1. Plant Material
Accessions of primarily wild perennial flax were obtained from the Germplasm Resources Information Network (GRIN) of the USDA-ARS, Plant Gene Resources of Canada (GRIN-CA), Kew Millennium Seed Bank, and several commercial sources. In total, 137 accessions were studied of L. alatum, L. altaicum Ledeb. ex Juz., L. aristatum Engelmann, L. austriacum, L. baicalense Juz., L. bienne, L. decumbens Desf., L. flavum L., L. grandiflorum, L. hirsutum L., L. hudsonoides Planch., L. leonii F.W.Schultz, L. lewisii, L. narbonense, L. pallescens Bunge, L. perenne, L. stelleroides, L. strictum, L. sulcatum Riddell, L. virgultorum Boiss. & Heldr. ex Planch, and L. viscosum. Eight accessions of annual, domesticated L. usitatissimum were included as check lines (Table 1).
Initial FGI evaluations of perennial flax from 2005–2008 occurred in a randomized common garden containing Linum altaicum, L. austriacum, L. baicalense, L. bienne, L. campanulatum L., L. flavum, L. hirsutum, L. lewisii, L. perenne, L. sulcatum, L. tauricum Willd., L. tenuifolium L., and L. thracicum Degen (N. Anderson and K. Betts, unpub. data). These early generations of seed were open-pollinated (OP), so the current species composition of these populations is unknown and may include interspecific hybrids [25,26]. Plants which survived the 2005 winter were selected for yield in 2006 and 2007 and replanted in 2008. Remnant seed from 2008 was planted in spring 2017, out of which the most vigorous plants were visually selected and used to start an “elite restart” nursery in fall 2017 (K. Betts, pers. comm., 2020–2021). In fall 2018, selections for CF and OS traits were made from the 2018 elite restart nursery, as well as from an additional small nursery containing open pollinated L. austriacum, L. lewisii, and L. perenne.
In the present study, the population ‘Selections-CF’ consists of 17 selections made in 2018 for the CF ideotype [1,2], based on growth habit, stem length, and flower diameter. The ‘Selections-OS’ population consists of 25 genotypes selected in 2018 for OS traits, primarily yield and seed size (1000 seed wt.) (Table 1). Within both populations, the top 9–10 parent genotypes were propagated as vegetative cuttings from field plants in fall 2018. Ten stem tip cuttings per genotype (>5 cm length) were harvested from the crown, labeled, sealed in bags [1.2 mL Get Reddi® Sandwich Bags, United States Plastic Corporation, Lima OH, USA], and put into a cooler on ice for transport to MN Ag. Exp. Station Plant Growth Facility, University of Minnesota (44°59′17.8′′ N, −93°10′51.6′′ W) before rooting. Cuttings were trimmed to 5–7 cm length using a sterile razor [GEM Carbon Steel Extra Sharp Single Edge Blade, The Razor Blade Co., Van Nuys, CA, USA], the lower leaves removed, and the cut stem base dipped into 1000 ppm Indole-3-butyric Acid (IBA), after which cuttings were inserted into pre-moistened foam propagation strips [ROOTCUBES® PLUS WEDGE®, Oasis Grower Solutions, Kent, OH, USA]. Cuttings were rooted for 5 wk. in a glass mist house (21/21 °C, day/night, 16 h; 0600–2200 h lighting with high pressure sodium high intensity discharge lamps or HIDs at a minimum set point of 150 μmol m−2 s−1 at plant level). An intermittent mist system, at a mist frequency of 10 min intervals (mist nozzles, reverse osmosis water) during 0600–2200 h with a 7 s duration was used. After rooting, cuttings were transplanted into 10.12 cm square deep pots [SVD-355-DEEP-BK-40, T.O. Plastics, Clearwater, MN, USA] filled with a soilless medium [Promix Mycorrhizae, Premier Horticulture Inc., Quakertown, PA, USA] and grown in a glass greenhouse at 16.7/15.5 °C day/night daily integral and a 16 h photoperiod (0600–2200 h; long days). Supplemental lighting was supplied during winter months and cloudy days by 400 w high pressure sodium high intensity discharge (HPS-HID) lamps, at a minimum of 150 μmol m−2 s−1 (plant level), with an 16 h photoperiod. These were grown as stock plants for cutting production in spring 2019. The OP seed from these clonal genotypes was also planted for evaluation in 2019 if n ≥ 59 seeds were available, in addition to the other selections.
2.2. Establishment of Common Garden Nursery
In spring 2019, all CF selections (17), OS selections (25), and species accessions (137) were grown in a common garden nursery in Rosemount, MN (44°42′58.2′′ N, −93°5′54.9′′ W) to compare phenotypic traits of interest within the same environment. Accessions were sown in 288 plug trays [Landmark Plastic, Akron, OH, USA] with soilless germination media [Berger BM2 Germination Mix, Berger, Saint-Modeste, QC, Canada] and covered with fine vermiculite [Palmetto Vermiculite Medium A-2, Palmetto Vermiculite, Woodruff, SC, USA] in wk. 14 and 15 (5, 12 April 2019). Due to the limited quantity of seed, four accessions (n = 59 seeds each) were planted by hand in each 288-plug tray, leaving an empty row between accessions to prevent cross contamination. For all breeding populations, n ≤ 288 seeds/genotype were sown using a vacuum seeder (E-Z Seeder, E-Z Seeder, Inc., Baraboo, WI, USA) into 288-plug trays. All plug trays were placed in a mist house for 4 h to moisten the soilless medium using an intermittent mist system (St. Paul MN Plant Growth Facility, University of Minnesota, 1552 Gortner Ave, St Paul, MN, USA) at a mist frequency of 10 min intervals (mist nozzles, reverse osmosis water) during 0600–2200 h with a 7 s duration (21/21 °C, day/night, 16 h; 0600–2200 h) with lighting supplied by high pressure sodium high intensity discharge (HID) lamps at a minimum set point of 150 μmol m−2 s−1. Once watered in, the trays were covered with plastic dome lids [Super Sprouter Standard Vented Humidity Dome 7”, Hawthorne Gardening Company, Vancouver, WA, USA] and transferred to a walk-in cooler for 2 wk. at 4/4 °C day/night in darkness to break seed dormancy (cold stratification), which is recommended for most wild Linum species (Barbara Atkins, STA laboratories, Longmont, CO, USA, pers. comm.). Trays were uncovered and misted by hand, as needed, over this 2-wk. period to maintain adequate moisture levels in the soilless medium. After the 2-wk. stratification, the dome lids were removed, and the trays were returned to the mist house for an additional 3 wk. Plug trays were then moved onto capillary mats in a greenhouse at 16.7/15.5 °C day/night daily integral and a 16 h photoperiod (0600–2200 h; long days) on wk. 19 and 20 (10, 17 May 2019). Supplemental lighting was supplied during cloudy days by 400 w high pressure sodium high intensity discharge (HPS-HID) lamps, at a minimum of 150 μmol m−2 s−1 at plant level. Fertigation (Mondays-Fridays) provided nutrients at a constant liquid feed (CLF) rate of 125 ppm N from water soluble 20-10-20 fertilizer. Accessions remained in the greenhouse until transplanting in wk. 24 (13 June 2019).
During wk. 15 and 16, 100 vegetative cuttings from the top CF and OS selections (indicated as ‘clone’; Table 1) were harvested to bulk up these genotypes for field trials. The propagation protocol was identical to the one outlined above, except these cuttings were sourced from greenhouse stock plants. In wk. 21, the rooted cuttings were moved to the identical greenhouse as the seedlings with the same fertigation regime; these also remained in the greenhouse until transplanting in wk. 24.
The common garden nursery was located at the Rosemount Research and Outreach Center, Rosemount, MN, USA. Accessions and seed propagated selections were randomized. Twenty seedlings and/or ten rooted cuttings per genotype were transplanted, with selection for early germination within genotypes. Planting spacing was 45.7 cm on-center within rows with 1.83 m row widths. The field was irrigated post-planting with 2.54 cm water. Irrigation continued throughout the summer to maintain a minimum of 2.54 cm water per week when there was insufficient rainfall. Weed control consisted of weekly mechanical tillage between rows, pre-emergent herbicide applications (Fortress®, OHP Inc., Bluffton, SC, USA) at the recommended rates, and bi-weekly hand weeding within rows.
2.2.1. Flowering and Survival Notes
Each individual plant in the nursery (seedling genotype, clonal ramet) was monitored weekly for flowering (≥1 open flowers). Flowering data collection began in wk. 27 and continued through wk. 43 for 17 wks. in total. The total number of weeks in flower was used to compare the flowering periods. Transplant survival was also recorded on a per-plant basis in wk. 43 at the end of the flowering season. Winter survival was recorded the following spring in wk. 19 (2020).
2.2.2. Measurements of Plant Size, Flower, Fruit, and Stem Characteristics
Given the size of the nursery (n = 2949 plants) it was not practical to carry out detailed phenotyping of every plant. Therefore, within each seed-propagated genotype, five plants were randomly selected for evaluation with the range of phenotypic variation for plant size, growth habit, and floral characteristics. For clonal selections, only three plants were selected per genotype due to genetic uniformity as clonal ramets.
Plant width was measured at the widest point (cm; width 1). The meter stick was rotated 90° from the width 1 axis to measure the shorter width (cm; width 2). Average widths/seed-propagated genotype or clonal selections were calculated.
Stem traits were recorded for three stems per plant and included the overall stem length (cm) measured from the crown to the stem apex, as well as the stem diameter (mm; recorded 30 cm from the apex of the stem). Stem lengths to the first basal branch > 5 cm (cm) in length were recorded as well as the proportion of unbranched stems. Both data points are useful selection criteria for both the CF ideotype and the yet-to-be-developed fiber ideotype.
Plant height was measured quantitatively as the distance from the soil line to the highest point on the plant without stretching out stems or holding up any part of the plant. A semi-ellipsoid volume, which combines all three size measurements into a geometrical shape most representative of the plant growth habit and is visualized as a dome with unequal horizontal axes, was calculated using the formula:
The base area of an ellipse was calculated to obtain an overall measure of plant width, irrespective of height, using the formula:
Circumference was calculated using the average width, according to the formula:
The height to width (h:w) ratio was calculated using the average width to determine whether plants were wider than they were tall, or vice versa, with the formula:
Finally, the eccentricity of an ellipse (e) was calculated to quantify the shape of each plant measured, using the formula:
Eccentricity of an ellipse (e) is a mathematical calculation with a range from 0–1 used to characterize the shape of a conical section. The eccentricity (e) of a perfect circle is zero. Thus, values of e approaching zero indicate a more circular shape, while values approaching one indicate an increasingly elliptical shape. In addition to geometry, eccentricity calculations are commonly applied in astronomy to describe the shape of an object’s orbit around another body [27]. However, the applications of eccentricity are far-ranging and have been used to characterize the shape of tumor cells in brain scans [28,29], yeast colonies [30], arterial stents [31], and the anomalous growth of tree rings following a landslide [32]. Cells of varying eccentricities have even been created to study the effect of cell shape on plasma membrane signaling [33]. In the present study, eccentricity values are used to estimate the shape of plant growth along the horizontal axis, as determined by measurements of width. Eccentricity values closer to zero, indicating more uniform growth, are desired.
Flower data were measured on three flowers per plant (reps.). Flower diameter (mm) and flower morph (classified as approach herkogamous [pin], reverse herkogamous [thrum], or homostylous) were recorded. Pin flowers are classified as having styles which are longer than the stamens, while thrum flowers are the reverse [22]. Homostylous flowers are those with styles and stamens of equal length. These data were used to estimate whether a population was monomorphic for one flower type or distylous (possessing both pin and thrum).
Flowers were also assigned ratings for petal overlap (1 ≥ 50% overlap, 2 ≤ 50% overlap) and flower shape (tube, funnel, or bowl) [17]. Tube-shaped flowers have a narrow corolla, often with overlapping petals, resulting in a small flower diameter. Funnel flowers are intermediate. Bowl flowers are those in which the corolla appears to open fully to almost at a 90° angle to the pedicel, resulting in the largest flower diameter possible [17]. For ornamental purposes, a bowl-shaped flower is desired to maximize flower diameter and showiness.
Flower color was determined for each seedling or clonal genotype, based on the Royal Horticultural Society (RHS) color charts [34]. The color was recorded for the distal end of the petal on full-sun days between the hours of 9 am and 3 pm. These color classifications were then converted to hexRGB values [35,36].
Dehiscence of the five bolls or capsules/plant were ranked for spontaneous capsule opening (shattering) on a 1–3 Likert scale for boll rating of each capsule where 1 = most dehiscent, 2 = intermediate, and 3 = least or non-dehiscent (most desirable) 0 capsules dehisced or nonshattering [1]. Mean scores/genotype and populations were calculated. Scoring is critical to enable selection for nonshattering bolls of pollinated plants [37], of particular importance for the oilseed harvest to prevent yield losses and preventing seed spread of CF and HP genotypes. Ideally, a rank of 3 (no boll dehiscence) is desirable in the ideotypes, although even annual flax has limited levels of shattering, despite its lengthy period of domestication [20]. Additionally, the number of underdeveloped seeds/capsule was recorded as a measurement of uniform seed development.
2.2.3. Statistical Analysis
The effect of population and genotype factors on the number of weeks in flower, width 1, width 2, height, semi-ellipsoid volume, base area, circumference, height to width ratio, eccentricity, flower diameter, number of weeks in flower, boll dehiscence, no. of underdeveloped seeds/capsule, stem length, stem length to 1st branch, proportion of stem unbranched, plant stature, plant width, and stem diameter were analyzed using independent, one-way Analysis of Variance (ANOVA) and mean separations (5% Tukey’s Honestly Significant Difference, HSD) using the Statistical Package for Social Sciences (SPSS, v. 25 for Windows, SPSS, Inc., Chicago, IL, USA). For population comparison analysis, phenotypic data were pooled by population. There was large variability in the sample size of each population as constrained by seed availability. To maintain statistical power, any species with n < 10 observations was dropped from the analysis. For analysis of genotypic differences, the chosen cutoff was n = 3 observations per genotype, which captured the majority of genotypes tested. Pearson correlations and descriptive statistics were calculated using SPSS to compare all traits analyzed by ANOVA. Flower morph was analyzed using a 1:1 Chi-square (χ2) test, as well as observations made in the present study. Petal overlap was analyzed using a 1:1 χ2. Flower shape was analyzed using a 1:1:1 χ2. Average summer and winter survival for each population were not compared using ANOVA since these amounted to one data point per population. Due to COVID-19 restrictions for travel to and working in the fields, plans to conduct the same phenotyping in Year 2 of growth (2020) were impossible to accomplish.
3. Results and Discussion
The main effects of population and genotype were very highly significant (p ≤ 0.001; Table 2) for all traits evaluated in the study (the no. weeks in flower, width 1, width 2, height, stem length, stem length to 1st branch, proportion of stems unbranched, stem diameter, flower diameter, height:width ratio, semi-ellipsoid volume, eccentricity, circumference, base area, plant stature, boll dehiscence, no. underdeveloped seeds/capsule). This significant variation within and among populations and genotypes of flax is not unexpected, given the diversity of sampling as well as inclusion of many wild species. Most importantly, significant genotypic differences provide for directional selection of traits for creation of CF, OS, and HP ideotypes.
3.1. Floral Characteristics
Traits relevant to CF potential for floral designing and HP potential in landscapes for “flower power” [38] include flower diameter (mm), weeks in flower, stem length (cm), and stem diameter (mm) (Figure 1). Since main effects are significant for all CF traits, mean separations of pooled groupings show differences among the wild species as well as OS and CF selections (Figure 1). Flower diameter, when pooled into species and ideotype groupings, was greatest in L. grandiflorum and L. lewisii, followed by CF, OS, L. altaicum, and L. austriacum (Figure 1A). Neither the CF or OS selection population had significantly different flower diameters compared to L. perenne, L. austriacum, L. altaicum, and L. hirsutum, and both selections had significantly larger flowers compared to L. baicalense, L. bienne, L. pallescens, and L. usitatissimum. Flower diameter differed greatly among populations on a genotype mean basis; the smallest diameter flowers (8–15 mm) occurred in L. bienne and L. usitatissimum (Figure 2). The significantly largest flowers occurred in three CF accessions (R-2.2.2DT-clone, S-294-3DT-clone, S-272-5DT-clone), two L. lewisii (Ames 33354, Ames 31364), and one OS genotype (S-293-5DT clone; Figure 2). On a genotype mean basis, a large range of flower diameters is observed for both selection populations, ranging from ~20 to −30 mm; L. bienne and L. usitatissimum were comparable in size and significantly smaller than any of the other populations (Figure 2). The CF selections alone exhibit a range of genotype mean values which encompasses all genotypes within populations of L. altaicum, L. austriacum, L. grandiflorum, L. hirsutum, L. lewisii, L. narbonense, and L. perenne. Such wide-ranging variation highlights the opportunities for selection within this population, as well as the challenge of achieving a consistent response to selection within a highly outcrossing species.
Given the shared evolutionary history of L. bienne and L. usitatissimum [5,39], flower size may not have undergone strong selective pressure over time as L. bienne gave rise to domesticated L. usitatissimum. Among breeding populations, while intentional selection for larger flower diameter had little impact overall with the CF selections’ mean flower diameter being slightly (non-significantly) greater than OS selections (Figure 1A). This may be partially due to CF populations being derived from a single cycle of selection in 2018, whereas three cycles of selection for OS traits have occurred since 2005. The existence of CF genotype means with significantly wider flowers than most OS types (Figure 2) allows for directed breeding and selection for CF improvement. Likewise, these CF genotypes would be useful in creating HP cultivars.
3.1.1. Flowering
A significantly greater number of weeks in flower was observed for the CF and OS selections compared to all wild species besides L. grandiflorum and L. austriacum (Figure 1B). The shortest flowering periods were observed for L. altaicum, L. baicalense, and L. lewisii, all of which were significantly less than the other populations; the remaining populations were intermediate to these extremes. As with flower diameter, L. usitatissimum and L. bienne exhibited similar responses in the number of weeks in flower. On a genotype mean basis, the range of flowering was widespread across all genotypes, ranging from 0 (non-flowering)–12.8 wks. (Figure 3). A lengthy ~13 wk. flowering period is unprecedented for HP that routinely flowering over a short period of time or reflowering after cutback or deadheading. This demonstrates the potential perennial flax has of becoming an annualized HP for the market [40]. Non-flowering types indicate the need for a cold treatment for Yr. 2 flowering unless these are bred for Yr. 1 blooming as annualized perennials [40]. Genotype mean differences also revealed a few key trends regarding the level of variation present within flax populations. Most notably, apart from two OS genotypes, all of the CF and OS selections exceeded the grand mean of 6.9 wk. in flower (Figure 3). This contrasts with L. austriacum, L. perenne, and L. bienne, which had genotypes in the 0–3 range, but also genotypes with >10 wk. in flower, on average. A distinct cluster of L. lewisii is observed on the low end of the range and all but two L. lewisii genotypes were below the grand mean (Figure 3).
3.1.2. Inflorescence and Stem Traits
The longest mean stem length was observed in L. pallescens (Figure 1C). This was significantly longer than all other populations besides L. lewisii, the CF and OS selections. As such, L. pallescens could be an excellent source for taller CF and HP types as well as for fiber. Overall, less variation was observed for stem length relative to other traits, with many populations exhibiting overlapping means, such as L. altaicum, L. austriacum, L. grandiflorum, L. hirsutum, L. lewisii, L. perenne, L. usitatissimum, and OS selections. Visual selection for improved stem length was evident for the CF selections, which had significantly longer stems compared to L. baicalense, L. bienne, L. grandiflorum, L. hirsutum, and L. usitatissimum.
Stem diameter is a complex trait affecting CF and fiber and, to a lesser extent, for OS and HP perennial flax. Large diameter stems are stronger, largely due to their greater cross-sectional area, and are, therefore, desirable for CF applications as well as HP types [41]. Several species in the present study stand out for their increased stem diameters relative to other populations. The largest stem diameters, on average, were observed for L. usitatissimum and were significantly greater than all other populations besides L. baicalense and L. grandiflorum (Figure 1D). These latter two populations were observed to have larger diameters compared to all except L. bienne. While L. pallescens had notably thin stems, it did not differ significantly from the majority of populations tested (Figure 1D). Both CF and OS populations were very similar for stem diameter, although OS selections were observed to have slightly thicker stems.
3.1.3. Petal Overlap
Petal overlap (“gappiness”) did not differ significantly from a 1:1 χ2 for L. altaicum, L. grandiflorum, L. pallescens, and OS selections (Table 3), possibly due to low sample sizes. In cases where petal overlap differed significantly from the 1:1 ratio, only L. bienne had a majority of flowers with <50% overlap. For populations with a majority of flowers with >50% overlap, the two highest test statistics were observed for CF selections and L. hirsutum. For L. austriacum and L. perenne, even though the majority of flowers had >50% overlap, both phenotypes were observed, and L. perenne was just under the threshold of significance. Most importantly, the improved ornamental quality of the CF selections relative to the OS selections is demonstrated.
3.1.4. Flower Shape
Bowl shaped flowers were the most frequently observed flower shape and all populations differed significantly from the 1:1:1 χ2 except for L. baicalense and L. bienne (Table 4). Linum bienne is the only species tested with an equal distribution of flower shapes. Overall, tube-shaped flowers were the rarest shape, which can generally be considered a positive finding, as such a tightly bound corolla would have little ornamental value. Tube shaped flowers occurred only in L. austriacum, L. bienne, L. usitatissimum, and CF selections [2]. Bowl shaped flowers, the most desirable ornamental phenotype, were generally the most common, especially among L. austriacum and OS selections. Other populations, including L. grandiflorum, L. perenne, and CF selections had a relatively large proportion of funnel-shaped flowers.
3.1.5. Flower Color
Flower color proportions for each Linum species and populations show distinct differences in color distributions for blue, red, and/or white tints, tones, and shades, based on RHS color codes and hexRGB values (Figure 4) [34,35,36]. The fewest number of flower color codes within a species/population occur in L. altaicum (Figure 4A) with 3 codes, and 4 codes occurring in L. usitatissimum (Figure 4J). Increasing numbers of colors range from 5 (L. baicalense, L. bienne, L. pallescens, Figure 4C,D,H, respectively) to even higher occurrences of flower color codes were in the remaining species and populations:, L. perenne L. austriacum, L. grandiflorum, L. hirsutum, L. lewisii, Selections–CF, and Selections–OS (Figure 4B,E–G,I,K,L). Flower color ratings were done by multiple, trained researchers. This, coupled with the large numbers of samples to evaluate prevented collection of flowers and placement in standard environmental conditions each day for ratings, meant that these ratings could have minor levels of variation in the tints (hue + white), tones (hue + grey) or shades (hue + black) categorization of each flower color, while the hue would have remained the same. Nonetheless, these initial ratings demonstrate the wide range of variation for flower color present within and among the species and populations for use in future flower color breeding and enhancement of CF and HP ideotypes. Future flower color evaluations need to be conducted with smaller sample sizes to ensure their ratings can be accommodated in a standard light environment. Digital imaging for characterizing flower color, floral patterns and UV signals pre- and post-pollination, may further aid our understanding of plant-pollinator interactions [37,42,43] once a pollinator ideotype is developed for perennial flax.
3.1.6. Styly
The observed style morphs for most species matched previous reports, if applicable, e.g., L. bienne, L. pallescens, L. perenne, and L. usitatissimum (Table 5). Mismatches in observed vs. reported style morphs occurred for L. austriacum, L. grandiflorum, L. hirsutum and L. lewisii. The style morph for L. grandiflorum had been identified as polymorphic distylous (Table 5). However, only homostylous flowers were observed in the present study, but homostylous, pin, and thrum flowers have been detected (Figure 5). To the best of our knowledge, this is the first known report of a homostylous L. grandiflorum. Further research into the cross compatibility of the three flower morphs is needed to determine fecundity (seed set) among cross-compatible and cross-incompatible style and stamen length differences [21,23]. Similarly, L. hirsutum is reported to be polymorphic distylous, yet two of the studied individuals produced homostylous flowers, and the population overall supports a 1:1:1 distribution of flower morphs (Table 5).
The second style morph inconsistency was observed in L. lewisii. Multiple reports have identified that L. lewisii is monomorphic approach herkogamous, possessing pin flowers (Table 5). In dichotomous keys, style morph is one of the main distinguishing features between L. perenne and L. lewisii, which are otherwise almost identical in appearance [23]. For this reason, it was surprising to find that 14/46 observations in L. lewisii were ‘thrum’ flowers. This either means previous misidentification and/or questions the purity of L. lewisii germplasm examined herein which will be addressed in future molecular studies. Likewise, such misidentifications have necessitated phenotyping styly morph upon first flowering in every new genotype under evaluation to provide additional confirmation or questioning of species identity.
While the species background of the CF and OS selections is unknown, these populations appear to be polymorphic distylous (Table 5). Data did not deviate from a 1:1 χ2, and no homostylous flowers were observed.
Linum altaicum (distylous) and L. baicalense (distylous and homostylous; Table 5) have not been previously studied for style morphs. Self and cross pollination studies will be needed to determine linkage with SI (L. altaicum) and/or SC (L. baicalense). Additional germplasm needs to be screened before clear styly trait association of the species can be made.
3.2. Capsule Shattering, Seed Development
The range of Year 1 capsule levels of shattering (1–3 Likert scale; 1 = fully dehiscent to 3 = non-shattering) were in the 1.5–2.0 range, denoting semi-shattering occurring throughout (Figure 6A). There were significant differences with L. bienne, L. austriacum, and L. perenne being nearly or fully dehiscent. The least dehiscent wild species (1.75–2.0) were L. altaicum, L. baicalense, L. grandiflorum, L. hirsutum, L. lewisii, L. pallescens (Figure 6A) while cultivated annual flax, L. usitatissimum had the least amount of shattering (2.25; Figure 6A). This is not surprising, given this crop’s long history of domestication as an OS crop. Lack of shattering would likewise be important for the OS, CF, and HP ideotypes to prevent seed drop in cultivated fields and gardens as well as for floral designs (fresh or dried). Regardless of the ideotype, considerable breeding and selection efforts are necessary to prevent shattering.
All species and selection populations had a minimum of 1.25 underdeveloped seeds/capsule (Figure 6B); most means overlapped among species. The highest number occurred in L. pallescens with a mean of ~2.25 underdeveloped seeds/capsule (Figure 6B), making this species the most variable for uniform ripening or seed development. The lowest number of underdeveloped seeds/capsule were in L. bienne, L. grandiflorum and L. usitatissimum. Ideally, for OS seed production, uniform and early seed ripening would aid in reducing the time between the first and second seed harvests each season which is a high priority goal in the OS crop ideotype for perennial flax. Breeding and selection for this trait is imperative for continued OS crop development.
3.3. Plant Dimensions
The CF selections had width 1 dimensions that were significantly greater than all populations except for OS selections, which exhibited similar, though slightly smaller, width 1 values (Figure 7A). The OS selections were also significantly greater than L. altaicum, L. baicalense, L. bienne, L. hirsutum, L. lewisii, and L. usitatissimum for width 1. The greatest width 1 observed among wild species was in L. perenne and L. austriacum, which had comparable mean values that were both significantly greater than L. baicalense, L. hirsutum, and L. usitatissimum. The smallest width 1 belonged to L. baicalense (Figure 7A).
The CF selections were also significantly larger than all populations for width 2 except the OS selections, which overlapped in distribution (Figure 7B). Linum baicalense was observed to have a significantly smaller width 2 than most species, excluding only L. hirsutum and L. usitatissimum. Among the wild species, L. perenne had the greatest width 2 measurements, on average, being significantly greater than L. baicalense, L. hirsutum, L. lewisii, and L. usitatissimum (Figure 7B). Wider plants would be desirable for HP selections with equal widths 1 and 2 to creating spherical plant habits, rather than for either CF (upright) or OS types (funnel form or V-shaped for mechanical cutting) [1,3].
The greatest average height was observed in CF selections, followed by OS (Figure 7C). Both L. usitatissimum, the CF and OS selections were significantly taller than L. baicalense, L. bienne, L. hirsutum, and L. lewisii. The remaining species L. altaicum, L. austriacum, L. grandiflorum, L. pallescens, and L. perenne were all similar in height (Figure 7C). This trait would be most advantageous for CF and fiber types, rather than HP or OS.
The integration of all three plant size measurements (width 1, width 2, height) into the calculation of semi-ellipsoid volume (cm3) illustrates the large differences in the size of the populations tested (Figure 7D). With this measurement, the CF selections are more than double the overall size of all species populations, besides L. austriacum and L. perenne, and they are significantly larger in volume than all populations except for overlapping with OS (Figure 7D). The small plant volume of L. lewisii is surprising (Figure 7D), given its close phylogenetic relationship with L. perenne and L. austriacum [5,23]. One possibility is that the climate and soil type of Minnesota is unfavorable for this species, even though Minnesota lies on the eastern border of its native range [45]. Linum lewisii is often found in alpine regions, although it is also known to be a native component of prairie grasslands. The wide range of biomes included in its native range suggests that adapting L. lewisii to MN for the purposes of developing a native perennial crop may still be feasible, if a collection can be located from a similar region as the target environment.
The OS selections have similarly large semi-ellipsoid volumes (cm3), at nearly twice the average size of L. grandiflorum; significantly larger than all populations besides L. austriacum and L. perenne (Figure 7D). This highlights the significant impact that even 1–3 generations of selection can have on vigor and adaptation of perennial flax to a new environment and bodes well for making progress in HP and fiber ideotype selection.
A larger plant volume was expected for L. grandiflorum given its annual life cycle and reported ornamental value. While it did flower profusely throughout the season (Figure 3), its volume did not differ significantly compared to most of the other species studied (Figure 7D). This could indicate that the genotypes tested were poorly adapted to the local environment.
For evaluating ornamental value of potential HP selections, it is also informative to consider the plant base area and circumference, which exclude plant height from the calculation (Figure 8A,B). Wider plants would ensure soil line coverage in the landscape for HP types. In general, the same relative pattern as a semi-ellipsoid volume is observed except that L. lewisii and L. bienne appear larger, potentially indicating that their size comes more from their width than their height. As with volume, the largest base area and circumference is found with CF selections, which overlap with OS types, and both are significantly greater than all other populations (Figure 8A,B). It is also notable that the visual differences between populations appear greatest for semi-ellipsoid volume (Figure 7D) compared to base area and circumference (Figure 3B and Figure 8A), yet the circumference shows a greater number of significant mean separations between populations. For example, L. baicalense has a significantly smaller circumference compared to L. altaicum, L. austriacum, L. bienne, L. grandiflorum, L. pallescens, L. perenne, CF and OS selections. A similar outcome occurs in L. usitatissimum, which has a significantly smaller circumference and base area relative to L. austriacum, L. grandiflorum, L. perenne, CF selections, but a semi-ellipsoid volume which only differs significantly from the CF and OS selection populations. For HP development, CF, OS, L. perenne, and L. austriacum would provide the greatest selection potential for both plant base area and circumference.
Ultimately, the choice of plant size measurement must relate to the breeding and selection goals of each ideotype. If the goal is to select for the largest or most compact plants overall, then semi-ellipsoid volume is superior, since it integrates all three size measurements. If the goal is a wide or narrow base, irrespective of height, then base area or circumference are the measurement(s) of choice. Base area may be a superior calculation criterion, as it integrates both widths 1 and 2, rather than using an average width to calculate circumference. However, based on this study, circumference creates greater statistical differences among populations.
Measurements characterizing plant shape may also prove useful for selecting for ornamental quality and growth vigor. The height to width ratio observed for L. usitatissimum was significantly larger than all other populations in the study, indicating that this species was much taller than it was wide (Figure 8C). In contrast, L. bienne grew much wider than it was tall, with a height to width ratio that was significantly less than L. altaicum, L. hirsutum, L. lewisii, L. pallescens, and L. usitatissimum. Anecdotally, L. bienne was often observed to have nearly horizontal growth, although a few genotypes did grow into an upright cushion shape. The height to width ratio of CF and OS selections was also low relative to the other populations tested, although these populations did not differ significantly from any others besides L. usitatissimum. Linum perenne is the wild species with the most similar height to width ratio as the selections. The remaining species had height to width ratios in the range of 0.6–0.8.
The goal of testing eccentricity is to determine which population has the most spherical footprint. This calculation is most relevant to HP uses where compact, ‘cushion’ shaped plants are generally the goal. Eccentricity values approaching zero indicate a nearly perfect sphere, while values approaching one indicate an increasingly elliptic shape. The most spherical, on average, out of all the populations were the CF selections, which had significantly lower eccentricity values than L. baicalense, L. hirsutum, L. lewisii, L. pallescens, and L. usitatissimum (Figure 8D). The same result occurs for the OS selections, except that this population did not differ significantly from L. hirsutum. Overall, in addition to being larger in size, both selection populations exhibited a more uniform and cushion shaped growth habit which would be desirable for the HP and bedding plant industry. Of all the wild species, L. perenne exhibited the most uniform growth habit with a mean eccentricity value of 0.5, which was significantly less than L. baicalense, L. lewisii, and L. pallescens. Of these latter three species, L. baicalense was the most elliptic, due to an average width 1 that was nearly double the average of its width 2.
Stem length to the first branch is an important trait for CF and OS production, enabling ease of stem harvest by hand (CF) or mechanically (OS). The shortest stems, which would be useful for bedding plant production of HP lines, as well as enhanced branching for HP production in cultivated settings, were found in L. bienne, L. grandiflorum, and L. usitatissimum (Figure 9A). The longer distance from the crown (soil line) to branching occurred in L. lewisii, which overlapped with L. baicalense, L. perenne, and CF/OS selections (Figure 9A). Thus, selection has been effective in both OS and CF types; similar development using other species will be beneficial.
The proportion of unbranched stems (length to first branch >5 cm/total stem length) were the highest in L. baicalense (~0.6) and significantly greater than all other species and selected populations (Figure 9B), indicating that this species has less basal branching than the others. Most other species and populations were ~0.5 with the significantly lowest proportions occurring in L. bienne and L. grandiflorum (Figure 9B). While branching is highly desirable in both HP and CF ideotypes, it depends on where it occurs (Figure 9A). Lower proportions of unbranched stems are desirable in the HP flax crop ideotype whereas higher proportions would be ideal for the CF ideotype. Crop development using L. bienne or L. grandiflorum will focus selection on higher branching capacity.
3.4. Plant Survival
The highest % summer survival (stand establishment to the end of the growing season) was observed for OS selections (93.2%; Table 6), followed by wild L. perenne (90.5%) and CF types (88.7%), which confirms that these selections readily adapt to transplanting with high stand establishment. Linum austriacum, L. pallescens, and L. altaicum also had 81.5%, 72.3% and 70.7% survival, respectively (Table 6). The lowest % summer survival was for annual flax, L. usitatissimum, at 24.4% (Table 6), which is a seed-propagated crop (direct seeded). This was not due to senescence, as plants were considered to have survived the summer through seed set. The most likely cause for this high rate of mortality may have been transplant shock, since it is direct seeded in commercial plantings [46]. Linum baicalense and L. grandiflorum also showed relatively poor summer survival, suggesting that significant effort would be required to adapt these species to transplanting. However, it would be interesting to study whether the same outcome was observed through direct seeding. For L. bienne, L. hirsutum, and L. lewisii summer survival was between 50–60% (Table 6), indicating that these are poorly adapted, but may be improved through several cycles of breeding.
Winter survival also varied greatly among the populations tested (Table 6). The greatest percent winter survival was observed for the OS selections (97.4%), which confirms that these genotypes are USDA Zone 4 (Z4) hardy and well adapted to MN winters. Linum perenne exhibited similar winter hardiness (94.3%), followed by CF selections at (91.5%; Table 6). The species L. altaicum, L. austriacum, and L. hirsutum all had winter survival > 70%, which suggests that they have the potential to be fully Z4 hardy following additional selection. Subsequent research has shown variation in species and CF/OS selections for winter survival comparisons of controlled freezing tests with field survival [24], indicating that perennial flax is selectable for CF, OS, HP and fiber ideotypes. The low winter hardiness observed for L. lewisii was surprising given its wide distribution over alpine and plains regions, which stretches as far north as Alaska and Canada [45]. This result may be due to the relatively poor vigor of the genotypes tested, as evidenced by their small semi-ellipsoid volume (Figure 7D) and poor summer survival (Table 6). Likewise, year to year differences in snow cover could have impacted the survival of the species. Of the species native to the steppes of Asia and Siberia [16], L. pallescens performed better than L. baicalense; the latter of which exhibited only 6.9% survival (Table 6), indicating that L. baicalense is only marginally hardy to USDA Z4. Despite its specific epithet, L. bienne behaved more like an annual in Z4, flowering and setting seed in the first year of growth and exhibiting only 0.5% winter survival (Table 6). As expected, the annual species L. grandiflorum and L. usitatissimum had 0% winter survival (Table 6).
3.5. Trait Correlations
The number of weeks in flower had highly significant positive correlations with several traits, including height, width 1, width 2, semi-ellipsoid volume, circumference, and base area (Table 7). Of these, the highest correlation coefficient observed was for width 2 (r = 0.432). Additionally, there was a significant (p ≤ 0.05) correlation with flower diameter. Weeks in flower showed highly significant negative correlation with eccentricity; significant (p ≤ 0.01) negative correlation with height to width ratio; significant (p ≤ 0.05) negative correlation with stem diameter (Table 7).
Not surprisingly, height showed a highly significant positive correlation with other size measurements, including width 1, width 2, height to width ratio, semi-ellipsoid volume, circumference, and base area (Table 7). Of these, the highest correlation coefficient was observed for semi-ellipsoid volume (r = 0.692). A highly significant positive correlation was also observed between height and stem length (r = 0.506). This correlation coefficient would be expected to increase as progress is made in selecting for upright growth habit. A significant (p ≤ 0.01) positive correlation between flower diameter and height was observed, as well as a highly significant negative correlation between height and eccentricity.
Similar to height, both width 1 and width 2 had highly significant positive correlations with all other plant size measurements, with the exception of height to width ratio and eccentricity, both of which displayed highly significant negative correlations (Table 7). The highest correlation coefficients observed for both widths 1 and 2 were for circumference (r = 0.969 and r = 0.968, respectively). This is not surprising, as the circumference measurement was calculated using the average of widths 1 and 2. Additionally, there were highly significant positive correlations between widths 1 and 2, stem length, and flower diameter; as well as negative correlations between widths 1 and 2 and stem diameter.
Height to width ratio had highly significant negative correlations with circumference and base area (Table 7), indicating that, as height increased, width also tended to decrease. There was also a significant (p ≤ 0.05) negative correlation with semi-ellipsoid volume. A significant (p ≤ 0.01) positive correlation was observed between height to width ratio and eccentricity, as well as a significant (p ≤ 0.05) positive correlation with stem length.
There was a highly significant negative correlation observed between semi-ellipsoid volume and eccentricity (Table 7), possibly due to larger selection populations, which also had the lowest eccentricity values. Semi-ellipsoid volume also displayed highly significant positive correlations with base area, circumference, and stem length, as well as a significant (p ≤ 0.05) positive correlation with flower diameter (Table 7). A significant (p ≤ 0.01) negative correlation between semi-ellipsoid volume and stem diameter was observed.
Highly significant negative correlations were found between eccentricity, base area, and circumference (Table 7), potentially for the same reasons as correlation of eccentricity and semi-ellipsoid volume. Interestingly, there was also a significant (p ≤ 0.05) positive correlation between stem diameter and eccentricity, indicating that more elliptic plants tended to have thicker stems.
Circumference had a highly positive correlation with base area (r = 0.970; Table 7), indicating that these are largely duplicative metrics for plant size irrespective of height. Both circumference and base area also showed highly significant positive correlations with stem length and flower diameter, and highly significant negative correlations with stem diameter. There was also a significant (p ≤ 0.01) negative correlation between stem diameter and flower diameter observed. Finally, stem length had a significant (p ≤ 0.05) negative correlation with stem diameter, and a highly significant positive correlation with flower diameter. As CF selection progresses, an even higher correlation between these traits should be expected. The degree of positive correlation between stem length and flower diameter could potentially be a good method of measuring progress towards the CF ideotype [1].
3.6. CF, OS Ideotype Modifications for Future Breeding
Based on these findings, we propose several changes to improve the CF and OS ideotype models [1,2,3,24]. Stem diameter measurements provided limited utility in the present study, but they should be included in future iterations of the ideotype model, as these data would be useful in selecting for fiber potential, which is a future product of interest for the FGI breeding program. It may be worthwhile to revisit the method of determining stem diameter to determine the best method for selection. In the present study, stem diameter was measured with calipers 30 cm below the stem apex, although several different methods of measuring stem diameter have been reported, such as photographing stem cross sections with an optical microscope and recording diameter using ImageJ [46,47]. While this method has higher accuracy, but it would not be impractical for efficient screening large numbers of breeding lines. Alternative methods measured stem diameter at different heights: 2 cm [48] and 15 cm above the soil line [41] or midpoint in the stem [49,50]. Thus, standardization of stem diameter measurements in perennial flax and future research to determine the best and efficient method for large sample size measurements are in order.
As noted earlier, additional refinements of trait measurements, particularly for flower color, shattering, and underdeveloped seeds at uniform harvest dates are also in need of attention. It is recommended that the same suite of ideotype traits be measured in the future as a means of quantifying the effect of CF and OS selection on target and non-target traits over time. It should be noted that the traits chosen for measurement in this study are neither final nor exhaustive. Consistency across years should be the goal, but as breeding priorities change, or new data is generated, it may be necessary to further refine the ideotype models.
3.7. Ornamental Potential of Wild Flax Species
As noted by Cullis [14] and Fu [15], there is a lack of formal research characterizing the ornamental potential of wild flax species for CF and HP use. Aside from taxonomic descriptions [6], only one other study compared trait values of wild Linum species [18]. This study evaluated five perennial (L. austriacum, L. hirsutum, L. narbonense, L. perenne, L. thracicum) and six annual (L. angustifolium, L. bienne, L. hispanicum, L. crepitans, L. grandiflorum, L. pubescens) for a suite of traits. The results of Poliakova and Lyakh [18] are compared with the present study in cases where there was overlap between the traits and species tested, namely L. austriacum, L. hirsutum, L. perenne, L. bienne, and L. grandiflorum. Flower diameters observed herein differed slightly from those reported by [18], particularly for L. grandiflorum and L. hirsutum, but also for L. austriacum, L. perenne and, to a lesser extent, L. bienne. In general, the flower diameter measurements by Poliakova and Lyakh [18] exceed those reported in the present study (Figure 1A). Plant heights reported herein were lower (Figure 7C) while the plant width values were greater (Figure 7A,B) than those reported [18]. Most of the germplasm tested by Poliakova and Lyakh [18] was sourced from the N.I. Vavilov Research Institute of Plant Industry (VIR) and the All-Russian Research Institute for Flax (VNIIL), with the exception of L. austriacum and L. hirsutum, which were collected from their native range in the southern steppe of Ukraine. Several years of breeding work on L. grandiflorum at Zaporozhye National University to develop cultivars with different flower colors and shapes were also performed [18,51]. These studies also differed in the age of germplasm tested (three years of growth) in comparison with the present study for year one trait values. Altogether, these differences in accession origin, testing environment, and age of plants may explain differences among Poliakova and Lyakh [18] and the present study. Still, the results raise questions about the range of genetic and phenotypic diversity captured by wild flax collections at VIR, VNIIL, USDA-GRIN, and GRIN-CA. We anticipate examining this diversity with molecular analyses of single nucleotide polymorphisms (SNPs) in these flax species and populations.
The best wild species to incorporate into CF breeding varies, depending on the trait(s) of interest. For flower diameter and stem length, L. lewisii is the best overall (Figure 1A,C). However, the short flowering period of L. lewisii is a significant drawback, as stem yield would low if the plants only flower for 4 wk. throughout the summer (Figure 1B). Given these constraints, L. austriacum is the most promising species overall, as it has large flower and stem diameters (Figure 1A,D), the longest flowering time of any wild perennial (Figure 1B), and relatively long stem length (Figure 1C). As previously discussed, L. pallescens has some favorable CF traits, but it lacks adequate flower and stem diameters (Figure 1A,D). Linum grandiflorum also has CF potential, as it has large flowers, a long flowering time, acceptable stem lengths, and thick stems (Figure 1A–D). Selection among these species to enhance current CF genotypes with superior postharvest vase life [2] will continue to improve the domestication of CF perennial flax for the market.
3.8. HP Crop Ideotype: Modifications and Expansion
Based on the phenotypic measurements of flax species and selected populations for CF and OS traits in comparison with potential HP crop requirements, we propose the following changes to the garden performance or HP ideotype [1] for use in future directed breeding and selection. This ideotype will have increasing importance moving forward with the breeding program. The HP ideotype includes seven traits for the ideal phenotype: uniform habit, long flowering period, large flowers, vibrant color, >50% petal overlap, unique characteristics and pollinator services [1,3]. Selections of HP in advanced trials among the CF and OS selections and breeding lines were made for genotypes with enhanced flower power and robust growth compared to existing cultivars with potential market value (D. Tork, unpub. data, 2018–2020). In the present study, a variety of new selection traits were found to create uniform HP habits for landscape uses or we were able to expand the currently defined HP traits.
3.8.1. HP Uniform Habit
Based on plant base area and circumference (Figure 8A,B), wider plants provide soil line coverage; such dimensions were found in L. perenne, L. austriacum, CF and OS selections (Figure 8A,B). Nearly equal height:width ratios (Figure 8C), such as in L. perenne, and low eccentricity values for semi-ellipsoidal plants (approaching zero; Figure 8D) would be choice trait selections for larger-sized HP plants. The optimal height to width ratio depends largely on the selection objectives. For a compact bedding plant, a uniform cushion shape is desired, so the ideal height to width ratio would approximate 0.8–1.2 [52,53]. For a larger, bush type habit that appears more like a small HP shrub, a wider base may be acceptable to cover a larger footprint. Therefore, target height to width ratios might instead range from 0.5–0.7, similar to the observed values for the CF and OS selections. In contrast, CF or fiber types would ideally be much taller than they are wide, so values ranging from 1.0–2.0 would be ideal.
3.8.2. HP Long Flowering Period
Flowering times of up to ~13 weeks provide selection potential for extended flowering season of perennial flax (Figure 3), an unusual trait in herbaceous perennials [40]. Linum austriacum had the longest flowering time (Figure 1B), on average, of all the wild perennials tested, and its volume was second only to L. perenne (Figure 7D), indicating first year vigor. Although L. perenne had the fourth longest flowering time of any wild species, it grew wider and in a more uniform shape, compared to L. austriacum. Linum perenne also had improved winter survival compared to L. austriacum (Table 6). Both early and late flowering would be important for all HP flax as season extenders in the spring and fall periods. We have commenced selection for both seasons of flowering, ensuring frost tolerance in elite parents for future enhancement of HP flowering as ‘S-292-2-DT clone’ (Figure 3) illustrates.
3.8.3. HP Large Flowers
Large flower diameters are important for flower power and aesthetics in the landscape [38,40]. In the present study, the largest diameter flowers occurred in L. lewisii and L. austriacum, followed by L. altaicum, L. grandiflorum, and CF and OS selections, such as OS ‘S-293-5-DT-clone’ (Figure 1A and Figure 2). Accessions within these populations should be selected and intercrossed to enhance and fix large flower size for HP flax.
Whilst large flowers were identified initially in the HP ideotype [1], flower shape can be added in as an additional HP floral trait in the ideotype. Wild species and selection populations had varying occurrences of individuals within and among species and selection populations for flower shape categorizations of tube, funnel, or bowl [17]. Only L. austriacum and the CF/OS selection populations had genotypes with all three types of flower shapes whereas all other groups had either one or two representative classes (Table 4). If selecting for tubular flowers, L. bienne has the highest frequency. For funnel-shaped flowers, L. grandiflorum, L. perenne, and OS selections had the highest frequencies (Table 4) whereas for bowl-shaped flowers, the most likely sources would be L. austriacum, L. lewisii, L. perenne as well as the CF, OS selection populations. Thus, depending on the floral type of interest, sufficient genetic variation exists to directionally breed for any three flower shapes.
3.8.4. HP Vibrant Color
Flower color would be easily selected among and within species (Figure 4), depending on the floral color of interest. For most CF types and many HP selections, blue flowers would be advantageous as this color is rare among horticultural species and crops [3,24,38,40]. Selection of blue-flowered perennial flax [4] would be feasible in all of the wild species and CF/OS selections tested herein, with the exception of L. grandiflorum (Figure 4). Additional colors to create a complete color palette in a HP series could use L. grandiflorum for red and white flowers (Figure 4). White-flowered mutants have also been found in our populations of L. austriacum, L. lewisii, L. perenne and CF, OS selections [3,24]. Yellow flowers, while not found in the species tested herein, are known to occur in L. flavum, L. rigidum, and L. sulcatum [54]. Testing of this germplasm for yellow-flowered HP breeding and selection are underway.
3.8.5. HP > 50% Petal Overlap
Testing of petal overlap in the HP ideotype [1], showed that a lack of gappy flowers (>50% petal overlap), occurred in L. hirsutum and CF selection populations (Table 3). Genetic variation for this trait will ensure adequate selection potential within wild species and selections for future enhancement of this trait.
3.8.6. HP Unique Characteristics
Other unique traits of interest for HP product development noted in this study, include styly and non-shattering. To minimize or potentially prevent seed set and spread of perennial flax in the landscape or out of cultivation, homostyly would need to be selected against. Fortunately, among the tested CF and OS genotypes, none had homostyly (Table 5). However, this trait did appear in most other wild species, particularly in red-flowered L. grandiflorum, blue-flowered L. bienne and L. usitatissimum (Table 5). Desirable style morphs would be either pin or thrums, to minimize seed set at least within clonal cultivars or among cultivars with similar style morphs.
Non-shattering of seeds in HP would be desirable to prevent spread of any cultivars within cultivated settings during the flowering season. While L. usitatissimum had the highest rate of non-shattering (Figure 6A), it is an annual species which limits development of non-shattering HP types. Thus, selection within wild species or CF/OS selection populations would be required, followed by intercrossing and selection of nearly non-shattering types.
3.8.7. HP Pollinator Friendly
Information on honeybees’ attractiveness to annual flax (L. usitatissimum) is controversial, with some reporting it as “weak” [55] with honeybees collecting either solely pollen [37] or nectar [56], while others posit that it is more commonplace [57], particularly when a large number of colonies are near the production fields. While not evaluated per se, we observed multiple instances of pollinator visitation to annual and perennial flax accessions for nectar or pollen rewards, demonstrating the potential this trait has for ecosystem services of HP flax. It has yet to be established that honeybees visit perennial flax in a significant manner for pollination, especially if they’re grown for OS and/or fiber purposes. However, there are differences among annual and perennial flax species for pollination biology: annual flax is predominantly self-compatible and does not need pollinators whereas perennial species are predominantly distylous outcrossers (Table 5) requiring pollinator visitation for cross-pollination [58]. Annual flax only flowers 1x/season, prior to death, whereas perennial flax flowers throughout the growing season even with midseason and fall harvests with rapid regrowth and development of flowering shoots [3]. Thus, perennial flax may offer pollinators nectar/pollen rewards in times when other crops are nonflowering. This needs rigorous analyses to determine the types of pollinators, the degree of visitation and the material collected (nectar/pollen rewards).
In general, Linum austriacum or L. perenne would be considered the top perennial species for HP development although neither species is superior across all traits of interest. Thus, both should be considered in future breeding for HP cultivar development, especially if interspecific hybrids are female/male fertile. Linum grandiflorum may provide added value to the breeding program, as it had a long flowering time, unique flower colors, and a size and shape which is comparable with L. austriacum and L. perenne.
4. Conclusions
At least for the initial 1–3 generations of domestication, simultaneous selection for multiple traits can be achieved [3]. The most important outcome of the early stage domestication is increased vigor through adaptation to the local environment, as evidenced by increases in mean weeks in flower, plant size traits, and percent survival for both selection populations relative to wild species. Meanwhile, selection for OS and CF traits within the same population produced differential outcomes for traits of interest.
Eventually, it may be necessary to separate different ideotype populations into isolation nurseries which would be justified by referencing multi-year trends in ideotype trait values. For example, if gain from selection reaches a multi-year plateau for local adaptation traits, such as plant size or survival, then the combination of CF and OS selections in the same nursery could slow selection progress for traits of interest that are specific to each category. Likewise, if a trait that is important for one population begins to move in an unfavorable direction in another population over time, then there may be reason to separate the nurseries. For example, this could occur if selection for CF traits such as flower diameter or stem length began to negatively impact yield or vice versa. For now, in these early stages of selection, multiple ideotypes can be selected out of the same diverse pool of germplasm, which will conserve time, resources, and prevent genetic bottlenecks from occurring early in the domestication program. As progress towards the goal of perennial flax domestication advances, measurement of the ideotype traits should be conducted at a standard interval so that future trends in target and non-target traits can be recognized and responded to. As more data is collected, the practical application of these ideotype models could be greatly enhanced by the development of effective visualization tools to compare trends, trait correlations, and population differences over time. Therefore, continued utilization of this ideotype approach is not only an opportunity to study the process of crop domestication in real time, but also an actionable decision-making tool for domestication programs which need to balance multiple breeding objectives at once [3].
This study focused on the opportunity to improve Year 1 phenotypes. This was partly due to the interruptions caused by COVID-19, which prevented the same detailed phenotyping in Experiment 1 from occurring in 2020. We acknowledge that additional years’ data could alter the findings, although Year 1 data is demonstrative of the genetic variation present for the traits examined. Later it was noted that plants generally performed well in Year 2 and that Year 1 vigor was the area most in need of improvement (K. Betts, pers. comm., 2018).
For future breeders of perennial flax, the greatest future opportunities and questions involve examining trait changes across Years 1–3 of growth, determining the genetic basis for traits of interest, and finding effective ways of working around the high level of genetic diversity inherent in obligate outcrossers. There is also the opportunity to include fiber breeding program from the long-stemmed CF selections currently available. Finally, to fully realize the potential perennial flax, ecosystem services should be quantified, such as reductions in nutrient leaching, and the ability to uptake heavy metals for bioremediation projects [59,60,61,62,63]. Pollinator services could also be integrated into the breeding pipeline by measuring pollinator visitation, and identifying factors such as nectar production, flower color, or other traits that influence visitation [64,65]. The vast and varied opportunities for utilizing perennial flax make this a prime target for continued crop domestication efforts.
Author Contributions
Conceptualization, N.O.A., D.L.W. and D.G.T.; methodology, D.G.T. and K.J.B.; software, D.G.T.; validation, D.G.T., K.J.B. and N.O.A.; formal analysis, D.G.T.; investigation, D.G.T., N.O.A. and K.J.B.; resources, N.O.A. and D.L.W.; data curation, D.G.T.; writing—original draft preparation, D.G.T.; writing—review and editing, N.O.A.; visualization, D.G.T.; supervision, N.O.A. and D.L.W.; project administration, N.O.A.; funding acquisition, N.O.A. and D.L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Minnesota Agricultural Experiment Station, project number MAES-21-100; the Forever Green Initiative, grant numbers 00071139, CON000000084498; the USDA-ARS Crop Germplasm Committee grant number CON000000085489 for funding this research.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Sarah Gardner, Cora Rost, Michele Schermann, Cassandra Ward, Jessica Huege, and Rachel Schulz, all of whom assisted in the research for this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Domestication of perennial flax using an ideotype approach for oilseed, cut flower, and garden performance. Agronomy 2019, 9, 707. [Google Scholar] [CrossRef] [Green Version]
- Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Perennial flax: A potential cut flower crop. HortScience 2022, 57, 221–230. Available online: https://journals.ashs.org/hortsci/view/journals/hortsci/57/2/article-p221.xml (accessed on 12 August 2022). [CrossRef]
- Anderson, N.O.; Tork, D.G.; Hall, H.; Wyse, D.L.; Betts, K.J. Breeding perennial flax for ornamental and agronomic traits simultaneously during crop domestication increases the efficiency of selection. Acta Hort. 2022, in press.
- Bolsheva, N.L.; Melnikova, N.V.; Kirov, I.V.; Speranskaya, A.S.; Krinitsina, A.A.; Dmitriev, A.A.; Belenikin, M.S.; Krasnov, G.S.; Lakunina, V.A.; Snezhkina, A.V.; et al. Evolution of blue-flowered species of genus Linum based on high-throughput sequencing of ribosomal RNA genes. BMC Evol. Biol. 2017, 17, 253. [Google Scholar] [CrossRef] [Green Version]
- McDill, J.; Repplinger, M.; Simpson, B.B.; Kadereit, J.W. The phylogeny of Linum and Linaceae subfamily Linoideae, with implications for their systematics, biogeography, and evolution of heterostyly. Syst. Bot. 2009, 34, 386–405. [Google Scholar] [CrossRef]
- Ockendon, D.J. Taxonomy of the Linum perenne group in Europe. Watsonia 1971, 8, 205–235. [Google Scholar]
- Ockendon, D.J.; Walters, S.M. Linum L. In Flora of Europea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 206–211. [Google Scholar]
- Planchon, J.E. Sur la famille des Linées. Lond. J. Bot. 1847, 6, 588–603. [Google Scholar]
- Planchon, J.E. Sur la famille des Linées. Lond. J. Bot. 1848, 7, 165–186, 473–501, 507–528. [Google Scholar]
- Rogers, C.M. Relationships of the North American species of Linum (flax). Bul. Torrey Bot. Club 1969, 96, 176–190. [Google Scholar] [CrossRef]
- Small, J.K. Linaceae. In North American Flora; The New York Botanical Garden: New York, NY, USA, 1907; pp. 67–87. [Google Scholar]
- Winkler, H. Linaceae. In Die Natürlichen Pflanzenfamilien; Engler, H.G.A., Prantl, K.A.E., Eds.; Engelmann: Leipzig, Germany, 1931; pp. 82–130. [Google Scholar]
- Yuzepchuk, S.V. Linaceae. In Flora of the U.S.S.R.; Shishkin, B.K., Bobrov, E., Eds.; Botanical Institute of the Academy of Sciences of the USSR: Leningrad, Russian, 1974; pp. 82–130. [Google Scholar]
- Cullis, C. Linum. In Wild Crop Relatives: Genomic and Breeding Resources: Oilseeds; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 177–189. [Google Scholar] [CrossRef]
- Fu, Y.-B. A molecular view of flax gene pool. In Genetics and Genomics of Linum, Plant Genetics and Genomics: Crops and Models; Cullis, C.A., Ed.; Springer Int. Pub.: New York, NY, USA, 2019; pp. 17–37. [Google Scholar] [CrossRef]
- Diederichsen, A. Ex situ collections of cultivated flax (Linum usitatissimum L.) and other species of the genus Linum L. Genet. Resour. Crop. Evol. 2007, 54, 661–678. [Google Scholar] [CrossRef]
- Diederichsen, A.; Richards, K. Cultivated flax and the genus Linum L.: Taxonomy and germplasm conservation. In Flax: The Genus Linum; Muir, A.D., Westcott, N.D., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 22–54. [Google Scholar]
- Poliakova, I.; Lyakh, V. Comparative characteristics of vegetative and generative sphere in plants of wild flax species. Int’l. J. Agric. Environ. Res. 2017, 3, 3743–3754. [Google Scholar]
- Eastman, W. The History of the Linseed Oil Industry in the United States; T.S. Denison & Company, Inc.: Minneapolis, MN, USA, 1968. [Google Scholar]
- Bemiller, J.N.; Whistler, R.L.; Barkalow, D.G.; Chen, C.C. Aloe, chia, flaxseed, okra, pysllium seed, quince seed, and tamarind gums. In Industrial Gums; Whistler, R.L., Bemiller, J.N., Eds.; Academic Press: New York, NY, USA, 1993; pp. 227–256. [Google Scholar]
- Ruiz-Martín, J.; Santos-Gally, R.; Escudero, M.; Midgley, J.J.; Pérez-Barrales, R.; Arroyo, J. Style polymorphism in Linum (Linaceae): A case of Mediterranean parallel evolution? Plant Biol. 2018, 20, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liedl, B.E.; Anderson, N.O. Reproductive barriers: Identification, uses, and circumvention. In Plant Breeding Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1993; pp. 11–154. [Google Scholar]
- Pendleton, R.L.; Kitchen, S.G.; Mudge, J.; McArthur, E.D. Origin of the flax cultivar ‘Appar’ and its position within the Linum perenne complex. Int’l. J. Plant Sci. 2008, 169, 445–453. [Google Scholar] [CrossRef]
- Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Controlled freezing studies as a corollary selection method for winterhardiness in perennial flax (Linum spp.). Crop. Sci. 2022, 62, 1734–1757. [Google Scholar] [CrossRef]
- Jhala, A.J.; Hall, L.M.; Hall, J.C. Potential hybridization of flax with weedy and wild relatives: An avenue for movement of engineered genes? Crop. Sci. 2008, 48, 825–840. [Google Scholar] [CrossRef] [Green Version]
- Seetharam, A. Interspecific hybridization in Linum. Euphytica 1972, 21, 489–495. [Google Scholar] [CrossRef]
- Limbach, M.A.; Turner, E.L. Exoplanet orbital eccentricity: Multiplicity relation and the solar system. Proc. Natl. Acad. Sci. USA 2015, 112, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.D.; Aalhus, J.L.; Mapiye, C.; Rolland, D.C.; Larsen, I.L.; Basarab, J.A.; Baron, V.S.; McAllister, T.A.; Block, H.C.; Uttaro, B.; et al. Effects of diets supplemented with sunflower or flax seeds on quality and fatty acid profile of hamburgers made with perirenal or subcutaneous fat. Meat Sci. 2015, 99, 123–131. [Google Scholar] [CrossRef]
- Szczepankiewicz, F.; van Westen, D.; Englund, E.; Westin, C.-F.; Ståhlberg, F.; Lätt, J.; Sundgren, P.C.; Nilsson, M. The link between diffusion MRI and tumor heterogeneity: Mapping cell eccentricity and density by diffusional variance decomposition (DIVIDE). NeuroImage 2016, 142, 522–532. [Google Scholar] [CrossRef] [Green Version]
- De Prado, E.G.; Rivas, E.-M.; de Silóniz, M.-I.; Diezma, B.; Barreiro, P.; Peinado, M.J. Quantitative analysis of morphological changes in yeast colonies growing on solid medium: The eccentricity and Fourier indices. Yeast 2014, 31, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-K.; Ko, T.-G.; Oh, S.; Kim, J.-S.; Kang, W.C.; Jeon, D.W.; Yang, J.Y.; Choi, D.; Hong, M.-K.; Ahn, T.; et al. Comparisons of the effects of stent eccentricity on the neointimal hyperplasia between sirolimus-eluting stent versus paclitaxel-eluting stent. Yonsei Med. J. 2010, 51, 823–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šilhán, K. Tree-ring eccentricity in the dendrogeomorphic analysis of landslides—A comparative study. Catena 2019, 174, 1–10. [Google Scholar] [CrossRef]
- Rangamani, P.; Lipshtat, A.; Azeloglu, E.U.; Calizo, R.C.; Hu, M.; Ghassemi, S.; Hone, S.; Scarlata, S.; Neves, S.R.; Iyengar, R. Decoding information in cell shape. Cell 2013, 154, 1356–1369. [Google Scholar] [CrossRef]
- The Royal Horticultural Society. RHS Colour Chart, 3rd ed.; RHS & Flower Council of Holland: London, UK; Leiden, The Netherlands, 1995. [Google Scholar]
- The Azalea Society of America. Royal Horticultural Society, UCL and RGB Colors, Gamma = 1.4. 2021. Available online: https://www.azaleas.org/color-systems/ (accessed on 10 April 2022).
- The Royal Horticultural Society. Colours of the Royal Horticultural Society Colour Charts in sRGB, CIE L*a*b* (CIE Lab) and CIE L*C*h* (CIE LCh) System (Unofficial Colorimetric Results). 2021. Available online: https://rhscf.orgfree.com/ (accessed on 10 April 2022).
- Alex, A.H. Pollination of some oilseed crops by honey bees. In Texas Agricultural Experiment Station Progress Report; Texas A&M University: Lubbock, TX, USA, 1957. [Google Scholar]
- Anderson, N.O. Flower Breeding and Genetics; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Wang, Z.; Hobson, N.; Galindo, L.; Zhu, S.; Shi, D.; McDill, J.; Yang, L.; Hawkins, S.; Neutelings, G.; Datla, R.; et al. The genome of flax (Linum usitatissimum) assembled de novo from short shotgun sequence reads. Plant J. 2012, 72, 461–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, H.F.; Anderson, N.O. Creation of new floral products. Annualization of perennials—Horticultural and commercial significance. In Flower Breeding & Genetics: Issues, Challenges and Opportunities for the 21st Century; Anderson, N.O., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 49–64. [Google Scholar]
- Alcock, M.; Ahmed, S.; DuCharme, S.; Ulven, C. Influence of stem diameter on fiber diameter and the mechanical properties of technical flax fibers from linseed flax. Fibers 2018, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Chaki, J.; Dey, N. Applications of image color features. In Image Color Feature Extraction Techniques: Fundamentals and Applications, Briefs in Applied Science Technology; Chaki, J., Dey, N., Eds.; Springer: Singapore, 2021; pp. 73–83. [Google Scholar] [CrossRef]
- Garcia, J.E.; Greentree, A.D.; Shrestha, M.; Dorin, A.; Dyer, A.G. Flower colours through the lens: Quantitative measurement with visible and ultraviolet digital photography. PLoS ONE 2014, 9, e96646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, B.G. Floral biology and self incompatibility in Linum. Bot. Gaz. 1986, 147, 327–333. [Google Scholar] [CrossRef]
- Ogle, D.; St. John, L.; Peterson, S.J.; Tilley, D.J. Plant Guide for Blue Flax (Linum perenne) and Lewis flax (L. lewisii); USDA-Nat. Res. Cons. Ser. Idaho State Office: Boise, ID, USA, 2006. Available online: https://www.nrcs.usda.gov/Internet/FSE_PLANTMATERIALS/publications/idpmspg9388.pdf (accessed on 14 May 2022).
- Bourmaud, A.; Gibaud, M.; Baley, C. Impact of the seeding rate on flax stem stability and the mechanical properties of elementary fibres. Indus. Crops Prod. 2016, 80, 17–25. [Google Scholar] [CrossRef]
- Bourmaud, A.; Gibaud, M.L.; Lefeuvre, A.; Morvan, C.; Baley, C. Influence of the morphology characters of the stem on the lodging resistance of Marylin flax. Ind. Crops Prod. 2015, 66, 27–37. [Google Scholar] [CrossRef]
- Sharma, H.S.S.; Faughey, G.J. Comparison of subjective and objective methods to assess flax straw cultivars and fibre quality after dew-retting. Ann. Appl. Biol. 1999, 135, 495–501. [Google Scholar] [CrossRef]
- Couture, S.J.; DiTommaso, A.; Asbil, W.L.; Watson, A.K. Evaluation of fibre flax (Linum usitatissimum L.) performance under minimal and zero tillage in eastern Canada. J. Agron. Crop Sci. 2004, 190, 191–196. [Google Scholar] [CrossRef]
- Yang, F.; Liu, F.; Rowland, F. Effects of diurnal temperature range and seasonal temperature pattern on the agronomic traits of fibre flax (Linum usitatissimum L.). Can. J. Plant Sci. 2013, 93, 1249–1255. [Google Scholar] [CrossRef]
- Lyakh, V.A.; Soroka, A.I.; Boika, O.A. Diversity of ornamental flower traits and peculiarities of their inheritance in Linum grandiflorum Desf. Ind. Hort. J. 2018, 8, 106–110. [Google Scholar]
- Peter, A.; Thomas, P.; Van Iersel, M. Use of controlled water deficit to regulate poinsettia stem elongation. HortScience 2015, 50, 234–239. [Google Scholar] [CrossRef] [Green Version]
- Runkle, E.S. Height Control of Perennials: What Works and What Doesn’t. gpn: Greenhouse Product News. 2020. Available online: https://gpnmag.com/article/height-control-of-perennials-what-works-and-what-doesnt/ (accessed on 1 February 2022).
- USDA; NRCS. The PLANTS Database; National Plant Data Team: Greensboro, NC, USA, 2022. Available online: https://plants.usda.gov/home (accessed on 19 April 2022).
- Scullen, H.A.; Vansell, G.H. Nectar and pollen plants of Oregon. Oregon Agr. Expt. Sta. Bul. 1942, 412, 63. [Google Scholar]
- Smirnov, V.M. Cross-pollination of flax by bees. Pchelovodstvo 1954, 9, 53–55. (In Russian) [Google Scholar]
- Williams, I. The pollination of linseed and flax. Bee World 1988, 69, 145–152. [Google Scholar] [CrossRef]
- Duguid, S.D. Flax. In Handbook of Plant Breeding; Vollmann, J., Rajcan, I., Eds.; Springer Science + Business Media: New York, NY, USA, 2009; pp. 233–255. [Google Scholar]
- Angelova, V.; Ivanova, R.; Delibaltova, V.; Ivanov, K. Bio-accumulation and distribution of heavy metals in fibre crops (flax, cotton and hemp). Ind. Crops Prod. 2004, 19, 197–205. [Google Scholar] [CrossRef]
- Griga, M.; Bjelková, M. Flax (Linum usitatissimum L.) and Hemp (Cannabis sativa L.) as fibre crops for phytoextraction of heavy metals: Biological, agro-technological and economical point of view. In Plant-Based Remediation Processes, Soil Biology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 199–237. [Google Scholar] [CrossRef]
- Havel, L.; Trojan, V.; Krystofova, O.; Zitka, O.; Babula, P.; Zehnalek, Z. Phytoremediation potential of flax (Linum usitatissimum) to heavy metals with respect to biosynthesis of phytochelatins and activity of phytochelatin synthase. J. Biotech. 2010, 150, 223. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Hussain, S.; Kamran, M.; Chattha, M.S.; Ahmad, S.; Aqeel, M.; Rizwan, M.; Aljarba, N.H.; Alkahtani, S.; et al. Flax (Linum usitatissimum L.): A potential candidate for phytoremediation? Biological and economical points of view. Plants 2020, 9, 496. [Google Scholar] [CrossRef] [Green Version]
- Smykalova, I.; Vrbova, M.; Tejklova, E.; Vetrovcova, M.; Griga, M. Large scale screening of heavy metal tolerance in flax/linseed (Linum usitatissimum L.) tested in vitro. Ind. Crops Prod. 2010, 32, 527–533. [Google Scholar] [CrossRef]
- Jevtić, G.; Anđelković, B.; Radović, J.; Dinić, B.; Babić, S. Effect of alfalfa cultivar on pollinator visitation, seed yield and yield components. In Quantitative Traits Breeding for Multifunctional Grasslands and Turf; Sokolović, D., Huyghe, C., Radović, J., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2014; pp. 345–351. [Google Scholar] [CrossRef]
- Suso, M.J.; Bebeli, P.J.; Christmann, S.; Mateus, C.; Negri, V.; Pinheiro de Carvalho, M.A.A.; Torricelli, R.; Veloso, M.M. Enhancing legume ecosystem services through an understanding of plant–pollinator interplay. Front. Plant Sci. 2016, 7, 333. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one trait values related to the CF ideotype: (A) flower diameter (mm), (B) number of weeks in flower, (C) stem length (cm), and (D) stem diameter (mm). Mean separations (5% HSD) are displayed as letters above the columns denoting significance.
Figure 1.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one trait values related to the CF ideotype: (A) flower diameter (mm), (B) number of weeks in flower, (C) stem length (cm), and (D) stem diameter (mm). Mean separations (5% HSD) are displayed as letters above the columns denoting significance.

Figure 2.
Mean ± S.E. flower diameter (mm) for all wild species, CF and OS flax (Linum) genotypes with n ≥ 3 observations. Means separations (5% HSD) are displayed as letters above the columns denoting significance.
Figure 2.
Mean ± S.E. flower diameter (mm) for all wild species, CF and OS flax (Linum) genotypes with n ≥ 3 observations. Means separations (5% HSD) are displayed as letters above the columns denoting significance.

Figure 3.
Mean ± S.E. number of weeks in flower for all wild species, CF and OS flax (Linum) genotypes with n ≥ 3 observations. Means separations (5% HSD) are displayed as letters above the columns denoting significance.
Figure 3.
Mean ± S.E. number of weeks in flower for all wild species, CF and OS flax (Linum) genotypes with n ≥ 3 observations. Means separations (5% HSD) are displayed as letters above the columns denoting significance.

Figure 4.
Flower color (Royal Horticultural Society, RHS codes) proportion by Linum species and populations: (A) L. altaicum, (B) L. austriacum, (C) L. baicalense, (D) L. bienne, (E) L. grandiflorum, (F) L. hirsutum, (G) L. lewisii, (H) L. pallescens, (I) L. perenne, (J) L. usitatissimum, (K) Selections–CF, (L) Selections-OS. Number of observations are displayed in the center of each donut chart. RHS color codes were converted to hexRGB values [34,35,36].
Figure 4.
Flower color (Royal Horticultural Society, RHS codes) proportion by Linum species and populations: (A) L. altaicum, (B) L. austriacum, (C) L. baicalense, (D) L. bienne, (E) L. grandiflorum, (F) L. hirsutum, (G) L. lewisii, (H) L. pallescens, (I) L. perenne, (J) L. usitatissimum, (K) Selections–CF, (L) Selections-OS. Number of observations are displayed in the center of each donut chart. RHS color codes were converted to hexRGB values [34,35,36].

Figure 5.
Flower colors and floral morphs of Linum grandiflorum: (A) red and pink-flowered progeny of a red × white cross (white flowered type not shown) (B) From left to right: thrum (short-styled), homostylous (equal length), and pin (long-styled) flower morphs. Arrows indicate relative style (S) and anther (A) lengths. Note that sepals are partially removed in the two leftmost flowers to expose the floral organs.
Figure 5.
Flower colors and floral morphs of Linum grandiflorum: (A) red and pink-flowered progeny of a red × white cross (white flowered type not shown) (B) From left to right: thrum (short-styled), homostylous (equal length), and pin (long-styled) flower morphs. Arrows indicate relative style (S) and anther (A) lengths. Note that sepals are partially removed in the two leftmost flowers to expose the floral organs.

Figure 6.
Comparison of wild flax (Linum) species with CF and OS selections for mean ± S.E. Year 1 trait values related to oilseed ideotype: (A) boll rating (1–3; fully dehiscent to non-shattering) and (B) # underdeveloped seeds per capsule. Means separations (5% HSD): letters above columns.
Figure 6.
Comparison of wild flax (Linum) species with CF and OS selections for mean ± S.E. Year 1 trait values related to oilseed ideotype: (A) boll rating (1–3; fully dehiscent to non-shattering) and (B) # underdeveloped seeds per capsule. Means separations (5% HSD): letters above columns.

Figure 7.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one plant size trait values: (A) width 1 (cm), (B) width 2 (cm), (C) height (cm), and (D) semi-ellipsoid (S.E.) volume (m3). Means separations (5% HSD) are displayed as letters above the columns denoting significance.
Figure 7.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one plant size trait values: (A) width 1 (cm), (B) width 2 (cm), (C) height (cm), and (D) semi-ellipsoid (S.E.) volume (m3). Means separations (5% HSD) are displayed as letters above the columns denoting significance.

Figure 8.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one plant size and shape trait values: (A) base area (m2), (B) circumference (m), (C) height:width ratio, and (D) eccentricity (e). Mean separations are displayed as letters above the columns denoting significance.
Figure 8.
Comparison of wild flax species with CF and OS selections for mean ± S.E. year one plant size and shape trait values: (A) base area (m2), (B) circumference (m), (C) height:width ratio, and (D) eccentricity (e). Mean separations are displayed as letters above the columns denoting significance.

Figure 9.
Comparison of wild flax (Linum) species with CF and OS selections for mean ± S.E. Year 1 trait values related to the CF ideotype and would also be useful for future fiber length determinations: (A) length to first branch > 5 cm (cm) and (B) proportion of stem unbranched. Means separations (letters above the bars; 5% HSD) denote significance.
Figure 9.
Comparison of wild flax (Linum) species with CF and OS selections for mean ± S.E. Year 1 trait values related to the CF ideotype and would also be useful for future fiber length determinations: (A) length to first branch > 5 cm (cm) and (B) proportion of stem unbranched. Means separations (letters above the bars; 5% HSD) denote significance.


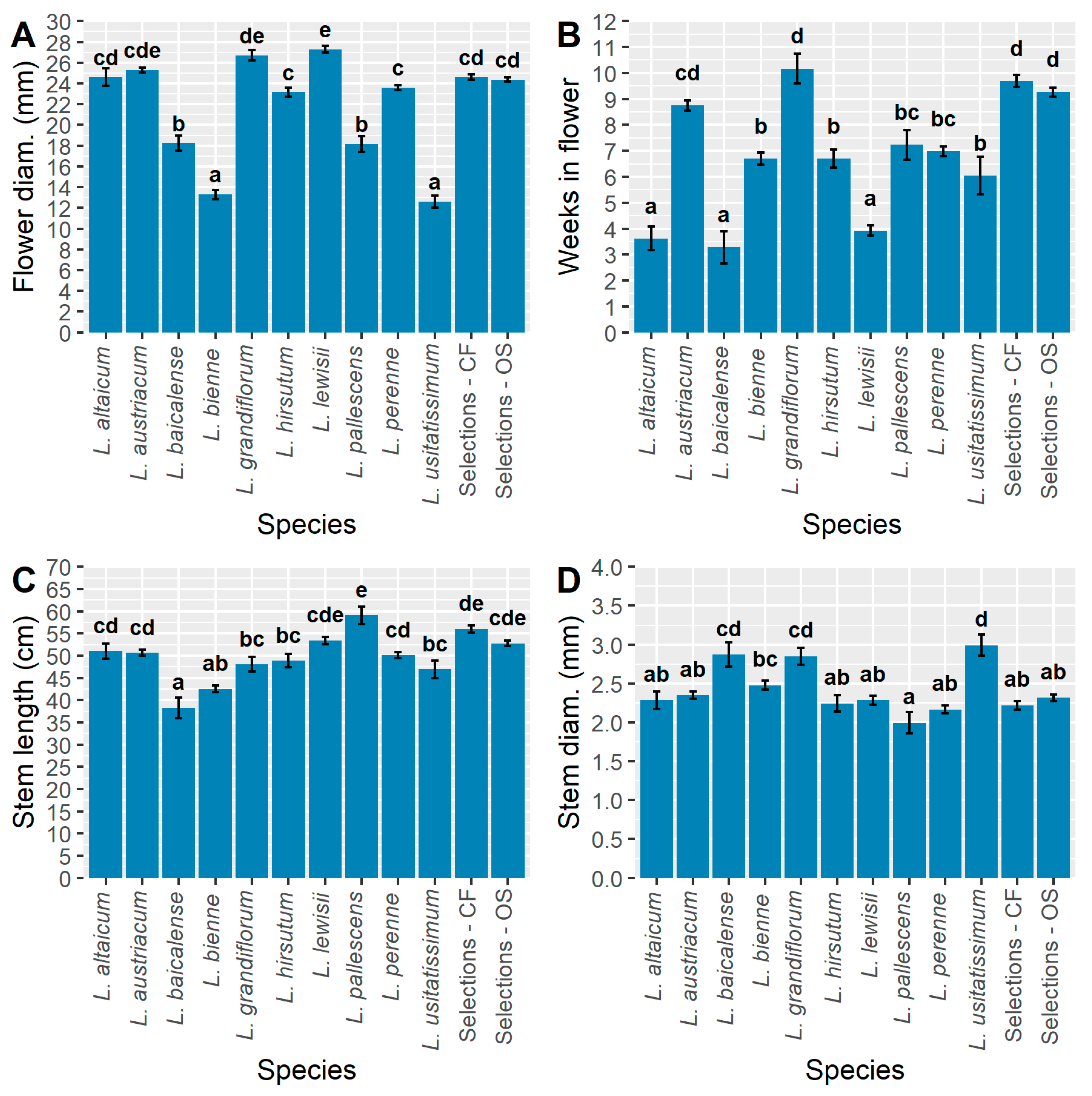{kind=link}
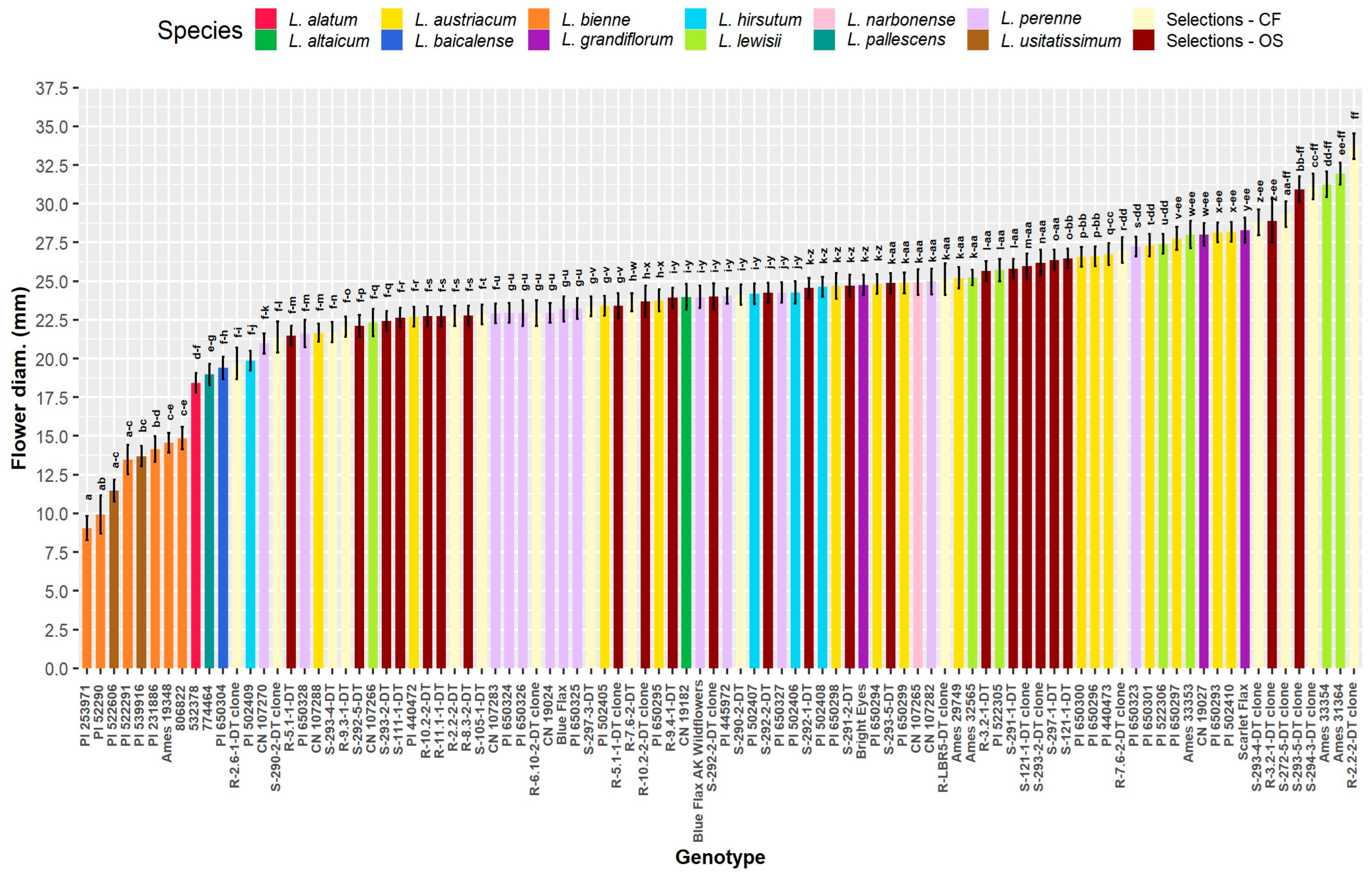{kind=link}
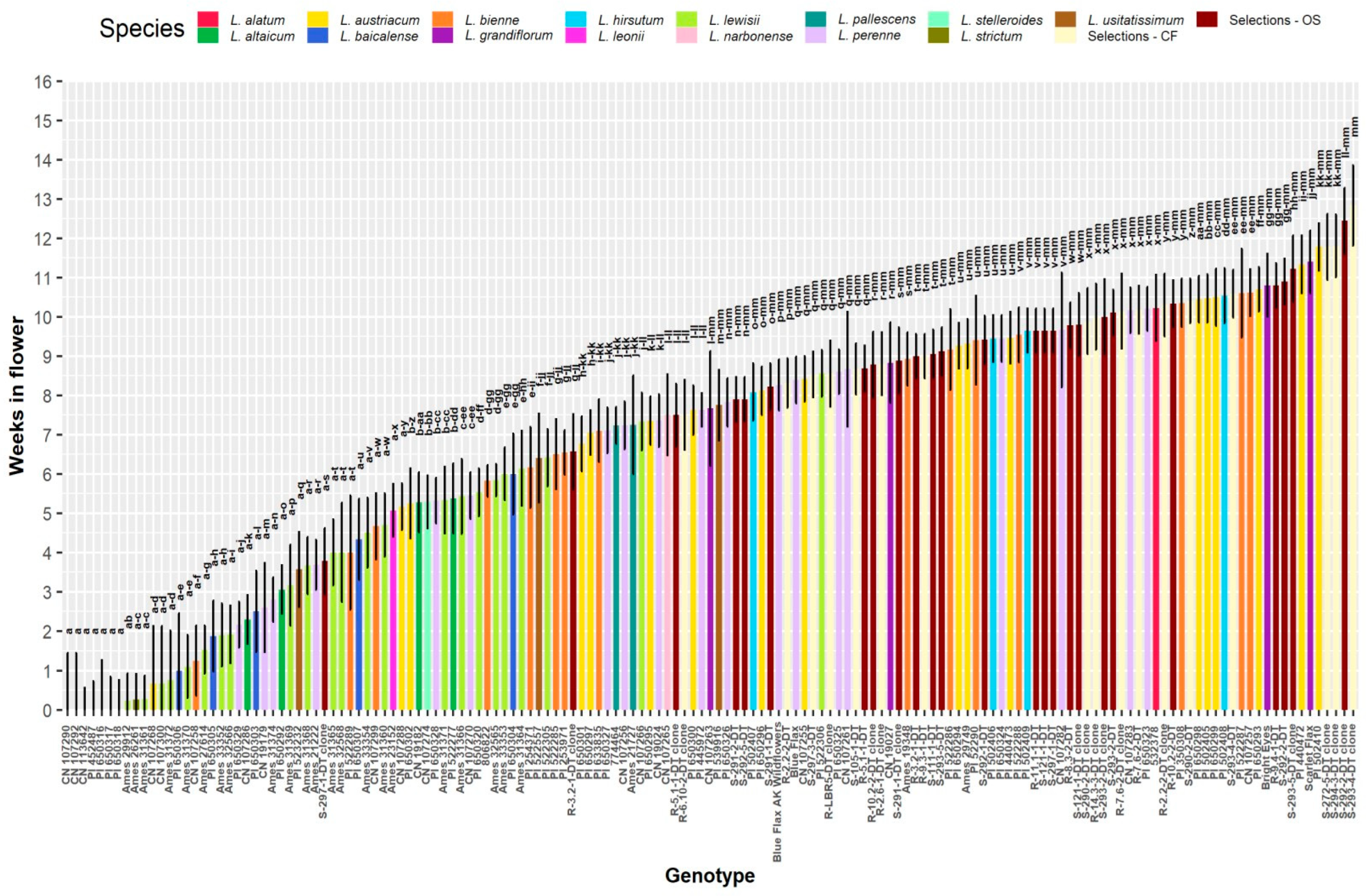{kind=link}
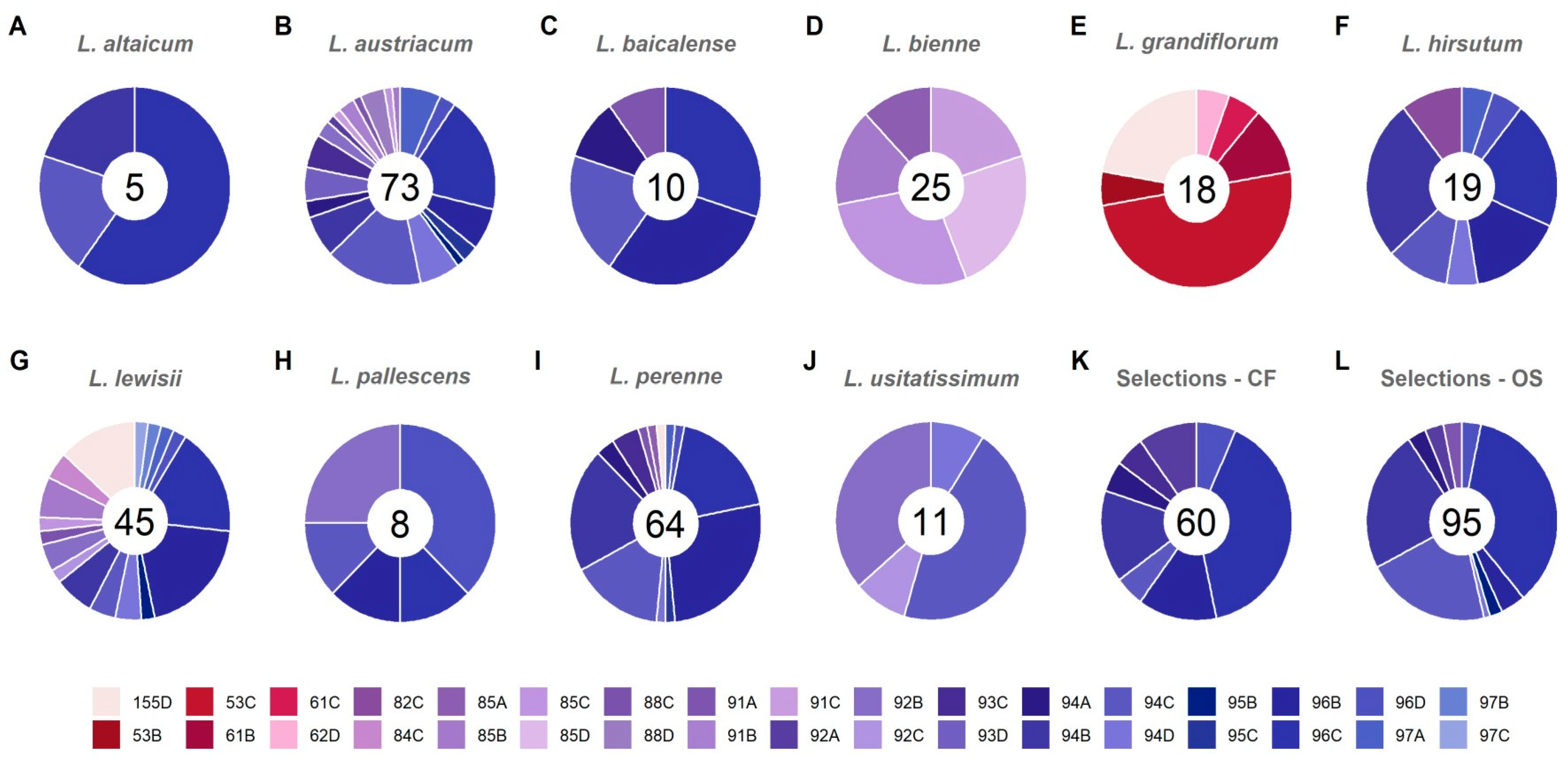{kind=link}
{kind=link}
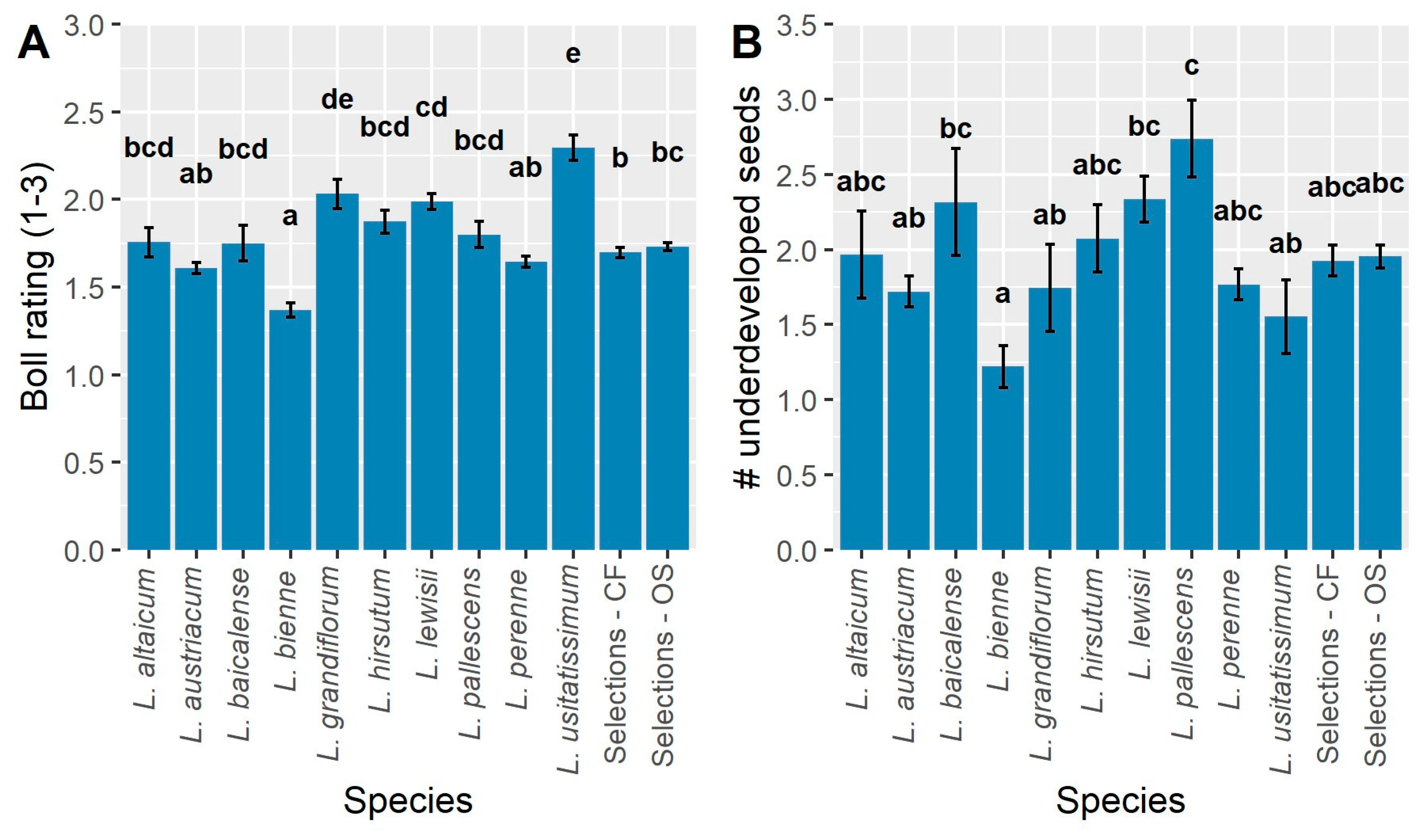{kind=link}
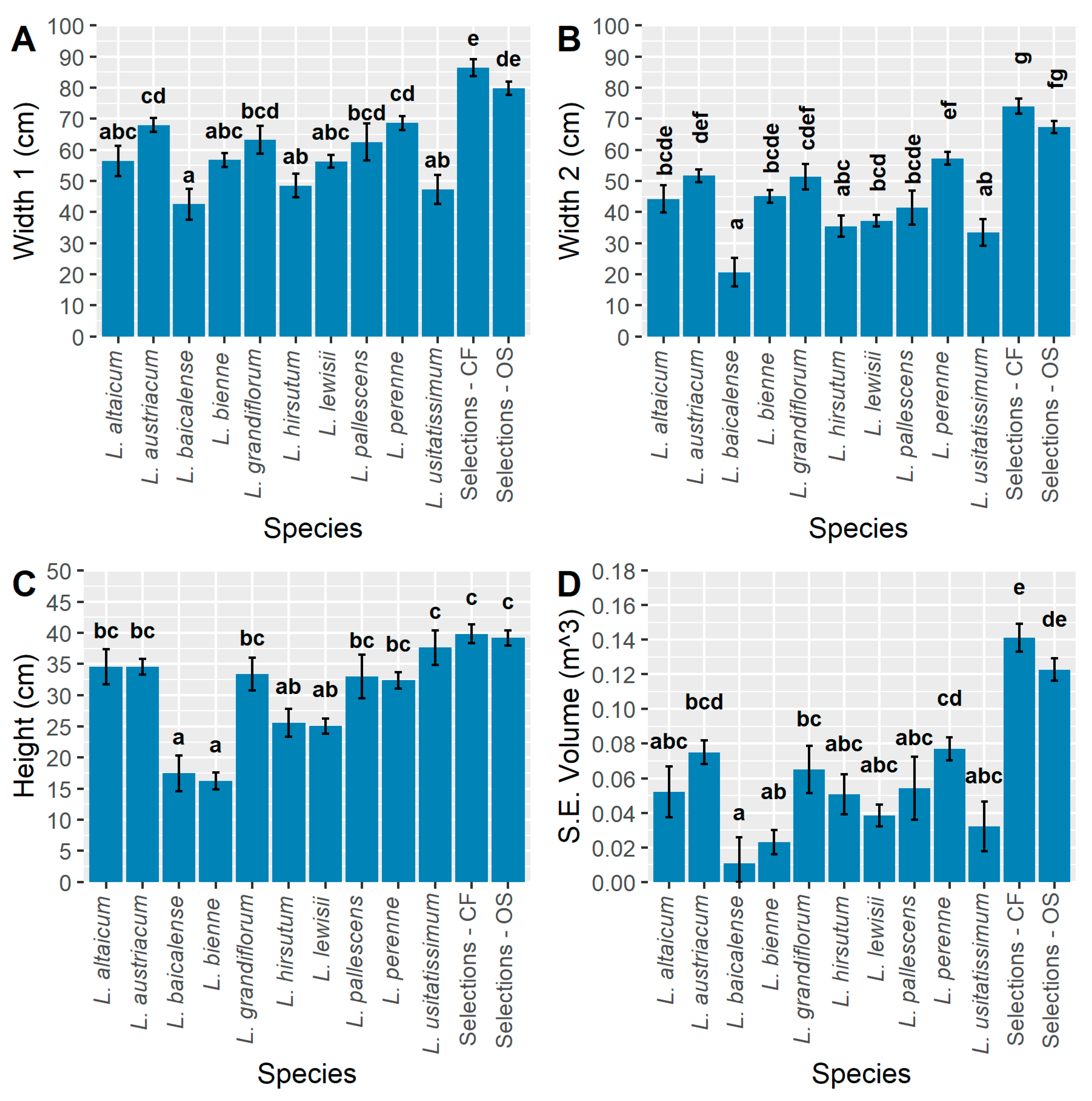{kind=link}
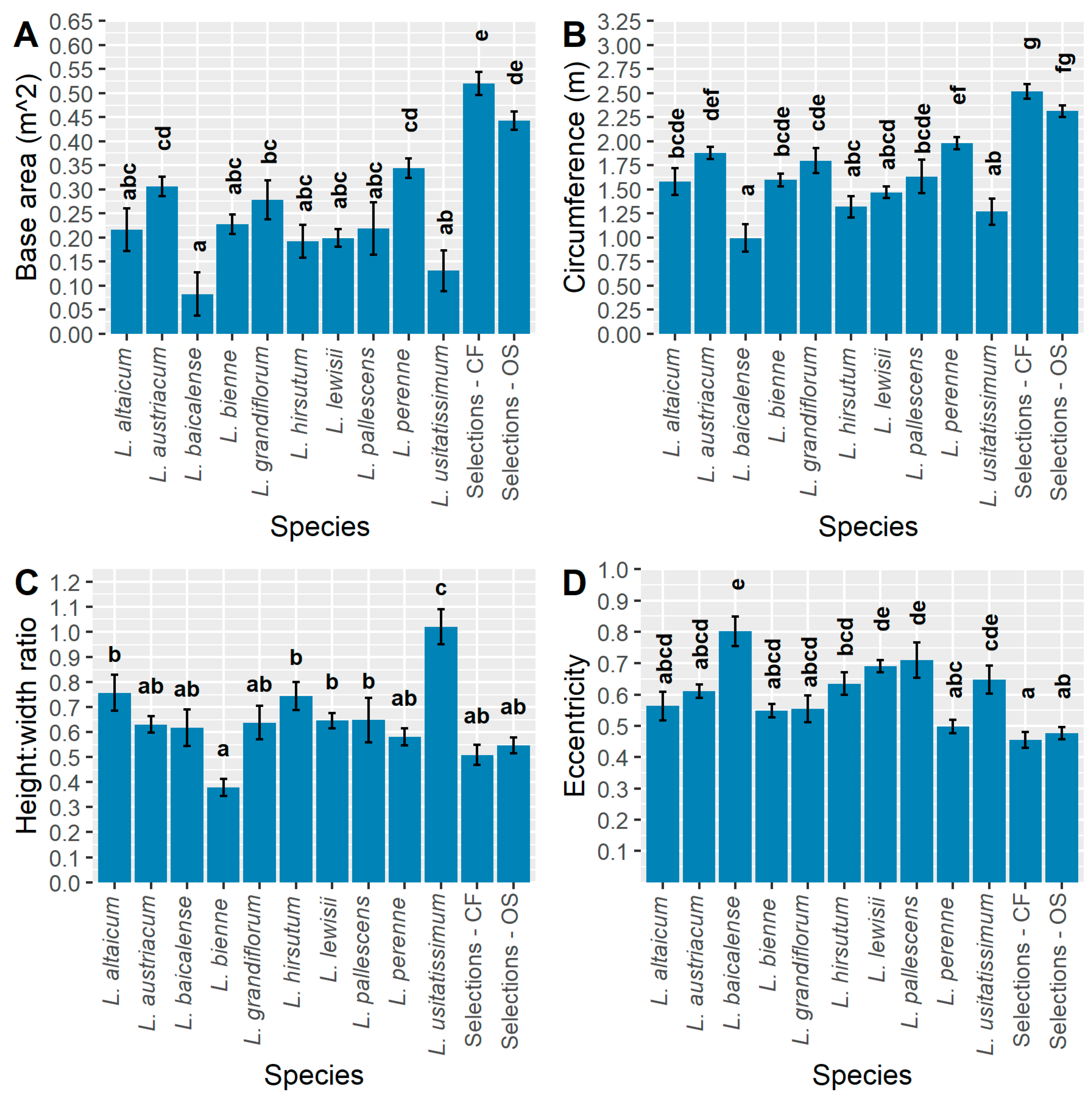{kind=link}
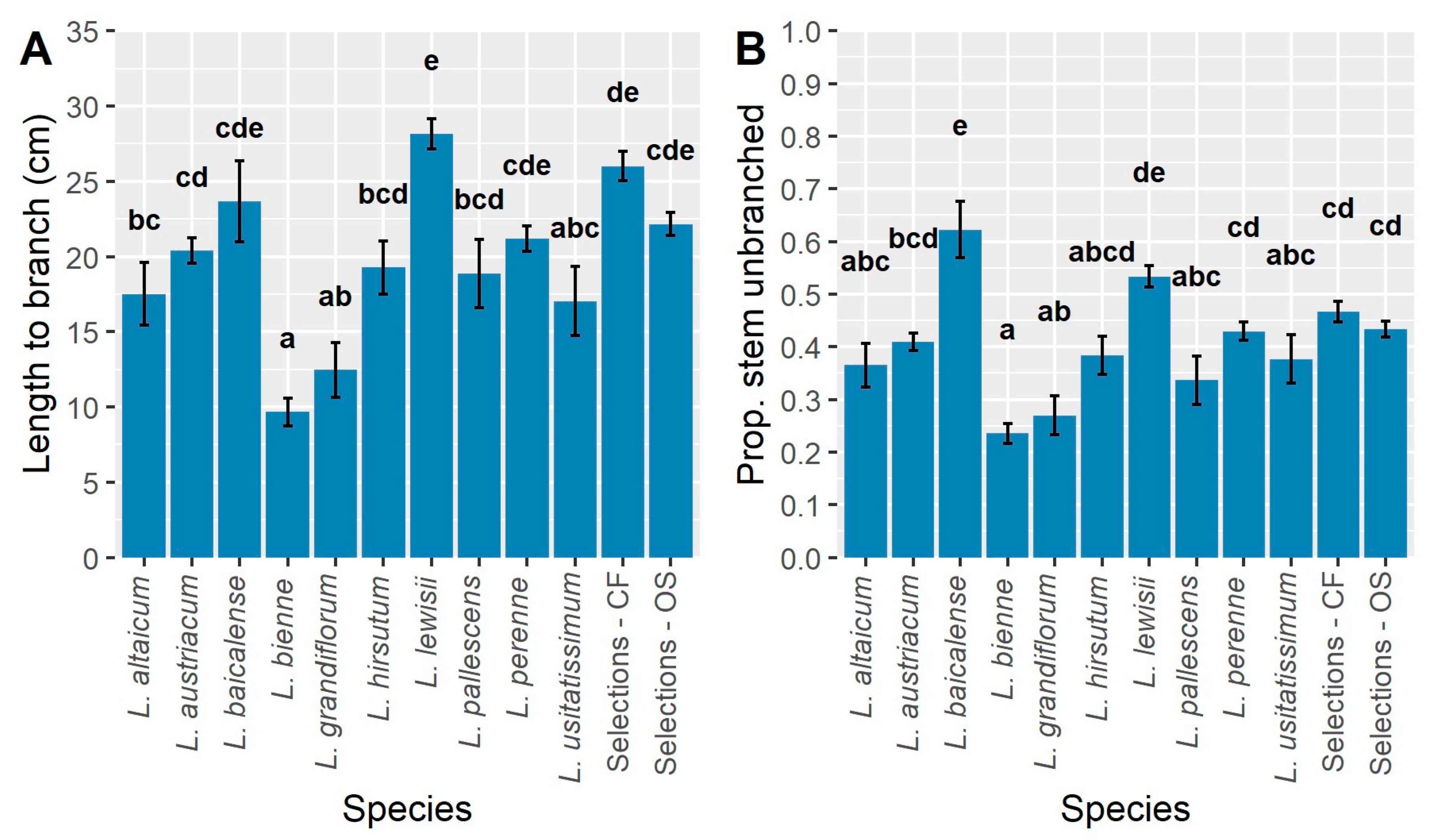{kind=link}
Table 1.
Linum species and populations (CFor OS selections), accession code, seed source, location and/or collection site of sources seeds used in the analysis.
Table 1.
Linum species and populations (CFor OS selections), accession code, seed source, location and/or collection site of sources seeds used in the analysis.
| Species | Accession | Seed Source, Location; (Collection Site) |
|---|---|---|
| L. alatum | 532378 | Kew Millennium Seed Bank, Royal Botanic Gardens, Wisley, Woking, Surrey, UK |
| L. altaicum | CN 107286 | GRIN-CA, Plant Gene Resources of Canada, Saskatoon, Canada |
| CN 19182 | GRIN-CA | |
| PI 522277 | USDA-GRIN, Ames, IA | |
| PI 650292 | USDA-GRIN (Baden-Wurttemberg, Germany) | |
| 202723 | Kew Millennium Seed Bank | |
| L. austriacum | Ames 29749 | USDA-GRIN (Ukraine) |
| CN 107255 | GRIN-CA | |
| CN 107268 | GRIN-CA | |
| CN 107288 | GRIN-CA | |
| PI 440472 | USDA-GRIN (Russian Federation) | |
| PI 440473 | USDA-GRIN (Russian Federation) | |
| PI 502405 | USDA-GRIN (Ukraine) | |
| PI 502410 | USDA-GRIN (Russian Federation) | |
| PI 650293 | USDA-GRIN (Hungary) | |
| PI 650294 | USDA-GRIN (Stavropol, Russian Federation) | |
| PI 650295 | USDA-GRIN (Poland) | |
| PI 650296 | USDA-GRIN (Tolbukhin, Bulgaria) | |
| PI 650297 | USDA-GRIN (Germany) | |
| PI 650298 | USDA-GRIN (Plovdiv, Bulgaria) | |
| PI 650299 | USDA-GRIN (Tolbukhin, Bulgaria) | |
| PI 650300 | USDA-GRIN (Bacs-Kiskun, Hungary) | |
| PI 650301 | USDA-GRIN (Armenia) | |
| PI 650302 | USDA-GRIN (Krym, Ukraine) | |
| L. baicalense | PI 650303 | USDA-GRIN (Mongolia) |
| PI 650304 | USDA-GRIN (Mongolia) | |
| PI 650305 | USDA-GRIN (Mongolia) | |
| PI 650306 | USDA-GRIN (Mongolia) | |
| PI 650307 | USDA-GRIN (Mongolia) | |
| L. bienne | 806822 | Kew Millennium Seed Bank |
| Ames 19348 | USDA-GRIN (Portugal) | |
| CN 107257 | GRIN-CA | |
| CN 107258 | GRIN-CA | |
| CN 107299 | GRIN-CA | |
| CN 113641 | GRIN-CA | |
| CN 113642 | GRIN-CA | |
| PI 231886 | USDA-GRIN (Belgium) | |
| PI 253971 | USDA-GRIN (Iraq) | |
| PI 254371 | USDA-GRIN (Delhi, India) | |
| PI 650308 | USDA-GRIN (Coimbra, Portugal) | |
| PI 522285 | USDA-GRIN (IA, USA) | |
| PI 522286 | USDA-GRIN (IA, USA) | |
| PI 522287 | USDA-GRIN (IA, USA) | |
| PI 522288 | USDA-GRIN (IA, USA) | |
| PI 522289 | USDA-GRIN (IA, USA) | |
| PI 522291 | USDA-GRIN (IA, USA) | |
| PI 522290 | USDA-GRIN (IA, USA) | |
| PI 633835 | USDA-GRIN (Delhi, India) | |
| L. decumbens | CN 19028 | GRIN-CA |
| L. flavum | CN 107281 | GRIN-CA |
| PI 650314 | USDA-GRIN (Germany) | |
| PI 650315 | USDA-GRIN (Denmark) | |
| L. grandiflorum | ‘Bright Eyes’ | Outsidepride Seed Source, LLC, 915 North Main Street, Independence, OR 97351 |
| CN 107259 | GRIN-CA | |
| CN 107260 | GRIN-CA | |
| CN 107263 | GRIN-CA | |
| CN 107287 | GRIN-CA | |
| CN 19026 | GRIN-CA | |
| CN 19027 | GRIN-CA | |
| ‘Scarlet Flax’ | Botanical Interests, Inc, 660 Compton Street, Broomfield, CO 80020 | |
| L. hirsutum | Ames 23759 | USDA-GRIN (Pest, Hungary) |
| CN 107271 | GRIN-CA | |
| PI 502406 | USDA-GRIN (Russian Federation) | |
| PI 502407 | USDA-GRIN (Russian Federation) | |
| PI 502408 | USDA-GRIN (Russian Federation) | |
| PI 502409 | USDA-GRIN (Russian Federation) | |
| PI 650316 | USDA-GRIN (Hungary) | |
| PI 650317 | USDA-GRIN (Poland) | |
| PI 650318 | USDA-GRIN (Romania) | |
| L. hudsonoides | 172989 | Kew Millennium Seed Bank |
| L. leonii | Ames 23152 | USDA-GRIN (Germany) |
| L. lewisii | ‘Maple Grove’ Ames 27614 | USDA-GRIN (UT, USA) |
| Ames 29912 | USDA-GRIN (USA, OR) | |
| Ames 31360 | USDA-GRIN (USA) | |
| Ames 31361 | USDA-GRIN (USA) | |
| Ames 31364 | USDA-GRIN (USA) | |
| Ames 31365 | USDA-GRIN (USA) | |
| Ames 31366 | USDA-GRIN (USA) | |
| Ames 31368 | USDA-GRIN (USA) | |
| Ames 31369 | USDA-GRIN (USA) | |
| Ames 31370 | USDA-GRIN (USA) | |
| Ames 31371 | USDA-GRIN (USA) | |
| Ames 31372 | USDA-GRIN (USA) | |
| Ames 32565 * | USDA-GRIN (USA) | |
| Ames 32566 | USDA-GRIN (USA) | |
| Ames 32568 | USDA-GRIN (USA) | |
| Ames 33352 | USDA-GRIN (NV, USA) | |
| Ames 33353 | USDA-GRIN (AZ, USA) | |
| Ames 33354 | USDA-GRIN (USA) | |
| CN 107266 * | GRIN-CA | |
| CN 107279 | GRIN-CA | |
| CN 107290 | GRIN-CA | |
| CN 107292 | GRIN-CA | |
| CN 107300 | GRIN-CA | |
| CN 19186 | GRIN-CA | |
| PI 452487 | USDA-GRIN (MT, USA) | |
| PI 522305 * | USDA-GRIN (IA, USA) | |
| PI 522306 * | USDA-GRIN (IA, USA) | |
| PI 650320 * | USDA-GRIN (UT, USA) | |
| L. narbonense | CN 107265 | GRIN-CA |
| L. pallescens | 774464 | Kew Millennium Seed Bank |
| Ames 21727 | USDA-GRIN (Asia) | |
| L. perenne | Ames 21222 | USDA-GRIN (Moldova) |
| Ames 31374 | USDA-GRIN (USA) | |
| Ames 32570 | USDA-GRIN (USA) | |
| ‘Blue Flax’ | Livingston Seed, 202 S Washington St, Norton, MA 02766 | |
| ‘Blue Flax’ | AK Wildflowers, Denali Seed Company, 6237 S Pere Marquette Highway, Pentwater, MI 49449 | |
| CN 107256 | GRIN-CA | |
| CN 107261 | GRIN-CA | |
| CN 107270 | GRIN-CA | |
| CN 107282 | GRIN-CA | |
| CN 107283 | GRIN-CA | |
| ‘Himmelszelt’ CN 19024 | GRIN-CA | |
| CN 19025 | GRIN-CA | |
| CN 19179 | GRIN-CA | |
| ‘Appar’ PI 445972 | USDA-GRIN (SD, USA) | |
| PI 650323 | USDA-GRIN (Hungary) | |
| PI 650324 | USDA-GRIN (Hungary) | |
| PI 650325 | USDA-GRIN (Hungary) | |
| PI 650326 | USDA-GRIN (Hungary) | |
| PI 650327 | USDA-GRIN (Hungary) | |
| PI 650328 | USDA-GRIN (ID, USA) | |
| PI 650329 | USDA-GRIN (ID, USA) | |
| L. stelleroides | CN 107274 | GRIN-CA |
| L. strictum | Ames 26261 | USDA-GRIN (Belgium) |
| PI 650331 | USDA-GRIN (Portugal) | |
| L. sulcatum | 354862 | Kew Millennium Seed Bank |
| L. usitatissimum | ‘Bison’ CN 33399 | GRIN-CA |
| ‘Linko de Riga’ CN 97295 | GRIN-CA | |
| ‘N.D. Resistant 52′ CN 97375 | GRIN-CA | |
| ‘Tammes #7 Pink’ CN 97427 | GRIN-CA | |
| ‘Novelty’ PI 522557 | USDA-GRIN (Ontario, Canada) | |
| ‘Bison’ PI 522606 | USDA-GRIN (ND, USA) | |
| ‘Raja’ PI 523232 | USDA-GRIN (Ontario, Canada) | |
| ‘Flanders’ PI 539916 | USDA-GRIN (Saskatchewan, Canada) | |
| L. virgultoum | CN 113624 | GRIN-CA |
| L. viscosum | 766777 | Kew Millennium Seed Bank |
| Selection Populations | ||
| Selections-CF | R-14.3-3-DT clone | Breeding program |
| R-2.2-2-DT | Breeding program | |
| R-2.2-2-DT clone | Breeding program | |
| R-2.6-1-DT clone | Breeding program | |
| R-6.10-2-DT clone | Breeding program | |
| R-7.6-2-DT | Breeding program | |
| R-7.6-2-DT clone | Breeding program | |
| R-9.3-1-DT | Breeding program | |
| R-LBR5-DT clone | Breeding program | |
| S-105-1-DT | Breeding program | |
| S-272-5-DT clone | Breeding program | |
| S-290-2-DT | Breeding program | |
| S-290-2-DT clone | Breeding program | |
| S-293-4-DT | Breeding program | |
| S-293-4-DT clone | Breeding program | |
| S-294-3-DT clone | Breeding program | |
| S-297-3-DT | Breeding program | |
| Selections-OS | R-10.2-2-DT | Breeding program |
| R-10.2-2-DT clone | Breeding program | |
| R-11.1-1-DT | Breeding program | |
| R-3.2-1-DT | Breeding program | |
| R-3.2-1-DT clone | Breeding program | |
| R-5.1-1-DT | Breeding program | |
| R-5.1-1-DT clone | Breeding program | |
| R-8.3-2-DT | Breeding program | |
| R-9.4-1-DT | Breeding program | |
| S-111-1-DT | Breeding program | |
| S-121-1-DT | Breeding program | |
| S-121-1-DT clone | Breeding program | |
| S-291-1-DT | Breeding program | |
| S-291-1-DT clone | Breeding program | |
| S-291-2-DT | Breeding program | |
| S-292-1-DT | Breeding program | |
| S-292-2-DT | Breeding program | |
| S-292-2-DT clone | Breeding program | |
| S-292-5-DT | Breeding program | |
| S-293-2-DT | Breeding program | |
| S-293-2-DT clone | Breeding program | |
| S-293-5-DT | Breeding program | |
| S-293-5-DT clone | Breeding program | |
| S-297-1-DT | Breeding program | |
| S-297-1-DT clone | Breeding program |
* Indicates ‘thrum’ flower types were observed for indicated L. lewisii genotype.
Table 2.
ANOVA (df, F ratio, Prob > F) for the effects of population (Linum species, selections) and genotype on the no. of weeks in flower, plant width 1 (cm), plant width 2 (cm), height (cm), stem length (cm), stem length to 1st branch (cm), proportion of stems unbranched, stem diameter (mm), flower diameter (mm), plant width (cm), height:width ratio, semi-ellipsoid volume (cm3), eccentricity, circumference (cm), base area (cm2), plant stature, boll dehiscence, and number of underdeveloped seeds/capsule.
Table 2.
ANOVA (df, F ratio, Prob > F) for the effects of population (Linum species, selections) and genotype on the no. of weeks in flower, plant width 1 (cm), plant width 2 (cm), height (cm), stem length (cm), stem length to 1st branch (cm), proportion of stems unbranched, stem diameter (mm), flower diameter (mm), plant width (cm), height:width ratio, semi-ellipsoid volume (cm3), eccentricity, circumference (cm), base area (cm2), plant stature, boll dehiscence, and number of underdeveloped seeds/capsule.
| Trait | Population | Genotype | ||||
|---|---|---|---|---|---|---|
| df | F Ratio | Prob > F | df | F Ratio | Prob > F | |
| No. weeks in flower | 11 | 66.743 | ≤0.001 | 150 | 19.996 | ≤0.001 |
| Plant width 1 (cm) | 11 | 19.019 | ≤0.001 | 148 | 6.938 | ≤0.001 |
| Plant width 2 (cm) | 11 | 29.230 | ≤0.001 | 148 | 7.956 | ≤0.001 |
| Height (cm) | 11 | 24.438 | ≤0.001 | 148 | 8.158 | ≤0.001 |
| Stem length (cm) | 11 | 20.078 | ≤0.001 | 129 | 13.763 | ≤0.001 |
| Stem length to 1st branch (cm) | 11 | 20.078 | ≤0.001 | 129 | 13.763 | ≤0.001 |
| Prop. of stems unbranched | 11 | 15.493 | ≤0.001 | 125 | 5.126 | ≤0.001 |
| Stem diameter (mm) | 11 | 7.930 | ≤0.001 | 123 | 3.126 | ≤0.001 |
| Flower diameter (mm) | 11 | 116.550 | ≤0.001 | 93 | 32.953 | ≤0.001 |
| Height:width ratio | 11 | 9.185 | ≤0.001 | 148 | 5.232 | ≤0.001 |
| Semi-ellipsoid volume (cm3) | 11 | 22.446 | ≤0.001 | 148 | 5.557 | ≤0.001 |
| Eccentricity | 11 | 12.401 | ≤0.001 | 148 | 2.114 | ≤0.001 |
| Circumference (cm) | 11 | 25.363 | ≤0.001 | 148 | 8.305 | ≤0.001 |
| Base area (cm2) | 11 | 22.114 | ≤0.001 | 148 | 6.454 | ≤0.001 |
| Plant stature | 11 | 12.564 | ≤0.001 | 148 | 4.550 | ≤0.001 |
| Boll dehiscence | 11 | 19.831 | ≤0.001 | 114 | 7.497 | ≤0.001 |
| No. underdev. sds./capsule | 11 | 4.629 | ≤0.001 | 114 | 3.222 | ≤0.001 |
Table 3.
1:1 Chi-square tests (1:1 χ2 test statistics, significance, df = 1) for petal overlap in flax species, CF and OS selection populations. For ornamental value in CF and HP genotypes, petal overlap > 50% (no gaps between petals) is desired to give the appearance of a larger, more full-looking corolla with “flower power”. Counts represent the number of individual plants within each category of petal overlap.
Table 3.
1:1 Chi-square tests (1:1 χ2 test statistics, significance, df = 1) for petal overlap in flax species, CF and OS selection populations. For ornamental value in CF and HP genotypes, petal overlap > 50% (no gaps between petals) is desired to give the appearance of a larger, more full-looking corolla with “flower power”. Counts represent the number of individual plants within each category of petal overlap.
| Population | Overlap > 50% | Overlap < 50% | 1:1 χ2 | Significance |
|---|---|---|---|---|
| L. altaicum | 4 | 1 | 1.80 | 0.180 ns |
| L. austriacum | 48 | 25 | 7.25 | 0.007 ** |
| L. baicalense | 9 | 0 | 9.00 | 0.003 ** |
| L. bienne | 6 | 19 | 6.76 | 0.009 ** |
| L. grandiflorum | 10 | 7 | 0.52 | 0.467 ns |
| L. hirsutum | 18 | 1 | 15.21 | ≤0.001 *** |
| L. lewisii | 33 | 12 | 9.80 | 0.002 ** |
| L. pallescens | 5 | 3 | 0.50 | 0.480 ns |
| L. perenne | 41 | 25 | 3.88 | 0.049 * |
| L. usitatissimum | 11 | 0 | 11.00 | ≤0.001 *** |
| Selections-CF | 46 | 14 | 17.07 | ≤0.001 *** |
| Selections-OS | 53 | 43 | 1.04 | 0.307 ns |
ns, *, **, *** denotes not significantly different, p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001, respectively.
Table 4.
Chi-square tests (1:1:1; test statistics, p-values, df = 2) for flax flower shapes in all populations and CF and OS selections tested. Flower shapes are classified as tube, funnel, or bowl [17]. Counts represent the number of individual plants within each category of plant shape. p-values of α ≤ 0.05 are highlighted in bold.
Table 4.
Chi-square tests (1:1:1; test statistics, p-values, df = 2) for flax flower shapes in all populations and CF and OS selections tested. Flower shapes are classified as tube, funnel, or bowl [17]. Counts represent the number of individual plants within each category of plant shape. p-values of α ≤ 0.05 are highlighted in bold.
| Population | Tube | Funnel | Bowl | Test Statistic | p-Value |
|---|---|---|---|---|---|
| L. altaicum | 0 | 0 | 5 | 10.00 | 0.007 |
| L. austriacum | 1 | 9 | 63 | 93.48 | ≤0.001 |
| L. baicalense | 0 | 3 | 6 | 6.00 | 0.050 |
| L. bienne | 6 | 9 | 10 | 1.04 | 0.595 |
| L. grandiflorum | 0 | 11 | 6 | 10.71 | 0.005 |
| L. hirsutum | 0 | 4 | 15 | 19.05 | ≤0.001 |
| L. lewisii | 0 | 2 | 43 | 78.53 | ≤0.001 |
| L. pallescens | 0 | 1 | 7 | 10.75 | 0.005 |
| L. perenne | 0 | 19 | 47 | 50.82 | ≤0.001 |
| L. usitatissimum | 3 | 8 | 0 | 8.91 | 0.012 |
| Selections-CF | 3 | 10 | 47 | 55.90 | ≤0.001 |
| Selections-OS | 0 | 13 | 83 | 124.56 | ≤0.001 |
Table 5.
Reported style morphs for flax species, field observations of style morphs (no. pin, no. thrum, no. homostylous plants), 1:1 (pin:thrum) or 1:1:1 (pin:thrum:homostylous) Chi-square (χ2) tests (df = 1 or 2, respectively), significance, and whether observations match previous reports of stylar condition (yes/no). Pin or approach herkogamous flowers are those in which the stigma protrudes above the anther height. Thrum or reverse herkogamous flowers have stigmas recessed below the anther height. Homostylous flowers are those in which the stigma and anthers are equal in height. Polymorphic species exhibit two (distylous) or three (tristylous) conditions within a single population and compatible interactions require flowers of different types to cross pollinate, promoting outcrossing. Monomorphic species exhibit one flower type only.
Table 5.
Reported style morphs for flax species, field observations of style morphs (no. pin, no. thrum, no. homostylous plants), 1:1 (pin:thrum) or 1:1:1 (pin:thrum:homostylous) Chi-square (χ2) tests (df = 1 or 2, respectively), significance, and whether observations match previous reports of stylar condition (yes/no). Pin or approach herkogamous flowers are those in which the stigma protrudes above the anther height. Thrum or reverse herkogamous flowers have stigmas recessed below the anther height. Homostylous flowers are those in which the stigma and anthers are equal in height. Polymorphic species exhibit two (distylous) or three (tristylous) conditions within a single population and compatible interactions require flowers of different types to cross pollinate, promoting outcrossing. Monomorphic species exhibit one flower type only.
| Population | Reported Style Morph | Reference | No. Pin | No. Thrum | No. Homostylous | χ2 Test | Test Statistic | Sig. | Match Reports (y/n) |
|---|---|---|---|---|---|---|---|---|---|
| L. altaicum | N/A | N/A | 2 | 4 | 0 | 1:1 | 0.667 | ns | N/A |
| L. austriacum | Polymorphic distylous | [21,44] | 31 | 41 | 1 | 1:1:1 | 35.62 | <0.001 | No |
| L. baicalense | N/A | N/A | 3 | 1 | 6 | 1:1:1 | 3.80 | ns | N/A |
| L. bienne | Monomorphic homostylous | [21,44] | 0 | 0 | 25 | Yes | |||
| L. grandiflorum | Polymorphic distylous | [21,44] | 0 | 0 | 17 | No | |||
| L. hirsutum | Polymorphic distylous | [21,44] | 8 | 9 | 2 | 1:1:1 | 4.53 | ns | No |
| L. lewisii | Monomorphic approach herkogamous (pin) | [21] | 32 | 14 | 0 | 1:1 | 7.04 | >0.01 | No |
| L. pallescens | Monomorphic homostylous | [21] | 0 | 0 | 8 | Yes | |||
| L. perenne | Polymorphic distylous | [21,44] | 39 | 27 | 0 | 1:1 | 2.18 | ns | Yes |
| L. usitatissimum | Monomorphic homostylous | [21,44] | 0 | 0 | 11 | Yes | |||
| Selections-CF * | N/A | N/A | 24 | 36 | 0 | 1:1 | 2.40 | ns | N/A |
| Selections-OS * | N/A | N/A | 47 | 49 | 0 | 1:1 | 0.04 | ns | N/A |
ns denotes not significant. * CF and OS selections of unknown species background were tested under the assumption that these populations were polymorphic distylous.
Table 6.
Percent summer survival and % winter survival for flax species, CF and OS selections investigated for domestication potential. Percent summer survival is defined as the total number of transplants established (stand establishment) and survived through the 1st growing season (Year 1). Using the number of plants that survived the growing season, % winter survival was calculated based on the number that survived the winter (Year 2) as either biennials or perennials.
Table 6.
Percent summer survival and % winter survival for flax species, CF and OS selections investigated for domestication potential. Percent summer survival is defined as the total number of transplants established (stand establishment) and survived through the 1st growing season (Year 1). Using the number of plants that survived the growing season, % winter survival was calculated based on the number that survived the winter (Year 2) as either biennials or perennials.
| Population | % Summer Survival | % Winter Survival |
|---|---|---|
| Linum altaicum | 70.7 | 71.7 |
| L. austriacum | 81.5 | 76.5 |
| L. baicalense | 29.0 | 6.9 |
| L. bienne | 53.7 | 0.5 |
| L. grandiflorum | 37.2 | 0.0 |
| L. hirsutum | 56.3 | 70.8 |
| L. lewisii | 57.7 | 66.1 |
| L. pallescens | 72.3 | 55.9 |
| L. perenne | 90.5 | 94.3 |
| L. usitatissimum | 24.4 | 0.0 |
| Selections-CF | 88.7 | 91.5 |
| Selections-OS | 93.2 | 97.4 |
Table 7.
Pearson correlations (r) for weeks in flower, height (cm), width 1 (cm), width 2 (cm), height:width ratio, semi-ellipsoid volume (cm3), eccentricity, circumference (cm), base area (cm2), stem length (cm), stem diameter (mm), flower diameter (mm) for flax populations of L. altaicum, L. austriacum, L. baicalense, L. bienne, L. grandiflorum, L. hirsutum, L. lewisii, L. pallescens, L. perenne, L. usitatissimum, CF and OS selections. Pearson correlations were calculated on a genotype mean basis for the average of three stems/flowers.
Table 7.
Pearson correlations (r) for weeks in flower, height (cm), width 1 (cm), width 2 (cm), height:width ratio, semi-ellipsoid volume (cm3), eccentricity, circumference (cm), base area (cm2), stem length (cm), stem diameter (mm), flower diameter (mm) for flax populations of L. altaicum, L. austriacum, L. baicalense, L. bienne, L. grandiflorum, L. hirsutum, L. lewisii, L. pallescens, L. perenne, L. usitatissimum, CF and OS selections. Pearson correlations were calculated on a genotype mean basis for the average of three stems/flowers.
| Trait | Weeks in Flower | Height (cm) | Width 1 (cm) | Width 2 (cm) | Height:Width | Semi-Ellipsoid vol. (cm3) | Eccentricity | Circum. (cm) | Base Area (cm2) | Stem Length (cm)a | Stem Dia. (mm)a |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Height (cm) | 0.373 *** | 1 | |||||||||
| Width 1 (cm) | 0.393 *** | 0.386 *** | 1 | ||||||||
| Width 2 (cm) | 0.432 *** | 0.408 *** | 0.876 *** | 1 | |||||||
| Height:width ratio | −0.136 ** | 0.467 *** | −0.474 *** | −0.431 *** | 1 | ||||||
| Semi-ellipsoid volume (cm3) | 0.343 *** | 0.692 *** | 0.754 *** | 0.775 *** | −0.084 * | 1 | |||||
| Eccentricity | −0.226 *** | −0.188 ** | −0.163 *** | −0.564 *** | 0.130 ** | −0.292 *** | 1 | ||||
| Circumference (cm) | 0.426 *** | 0.410 *** | 0.969 *** | 0.968 *** | −0.467 *** | 0.789 *** | −0.373 *** | 1 | |||
| Base area (cm2) | 0.349 *** | 0.388 *** | 0.925 *** | 0.954 ** | −0.399 *** | 0.843 *** | −0.378 *** | 0.970 *** | 1 | ||
| Stem length (cm)a | 0.039 | 0.506 *** | 0.506 *** | 0.402 *** | 0.092 * | 0.482 *** | 0.001 | 0.471 *** | 0.433 *** | 1 | |
| Stem diameter (mm)a | −0.106 * | −0.080 | −0.157 *** | −0.179 *** | 0.038 | −0.121 ** | 0.118 ** | −0.174 *** | −0.160 *** | −0.103 * | 1 |
| Flower diameter (mm)a | 0.107 * | 0.159 ** | 0.203 *** | 0.184 *** | 0.006 | 0.121 * | −0.080 | 0.201 *** | 0.164 ** | 0.227 *** | −0.168 ** |
*** Correlation is significant at the 0.001 level (2-tailed). ** Correlation is significant at the 0.01 level (2-tailed). * Correlation is significant at the 0.05 level (2-tailed).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Ideotype Selection of Perennial Flax (Linum spp.) for Herbaceous Plant Habit Traits. Agronomy 2022, 12, 3127. https://doi.org/10.3390/agronomy12123127
AMA Style
Tork DG, Anderson NO, Wyse DL, Betts KJ. Ideotype Selection of Perennial Flax (Linum spp.) for Herbaceous Plant Habit Traits. Agronomy. 2022; 12(12):3127. https://doi.org/10.3390/agronomy12123127
Chicago/Turabian StyleTork, David G., Neil O. Anderson, Donald L. Wyse, and Kevin J. Betts. 2022. "Ideotype Selection of Perennial Flax (Linum spp.) for Herbaceous Plant Habit Traits" Agronomy 12, no. 12: 3127. https://doi.org/10.3390/agronomy12123127
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.