Relationships between Foliar Fungal Endophyte Communities and Ecophysiological Traits of CAM and C3 Epiphytic Bromeliads in a Neotropical Rainforest


Abstract
:1. Introduction
2. Materials and Methods
2.1. Host Ecophysiological Traits
2.2. Fungal Cultures
2.3. DNA Extraction and Collections
2.4. Statistics
3. Results
3.1. Endophyte Abundance, Richness, and Diversity
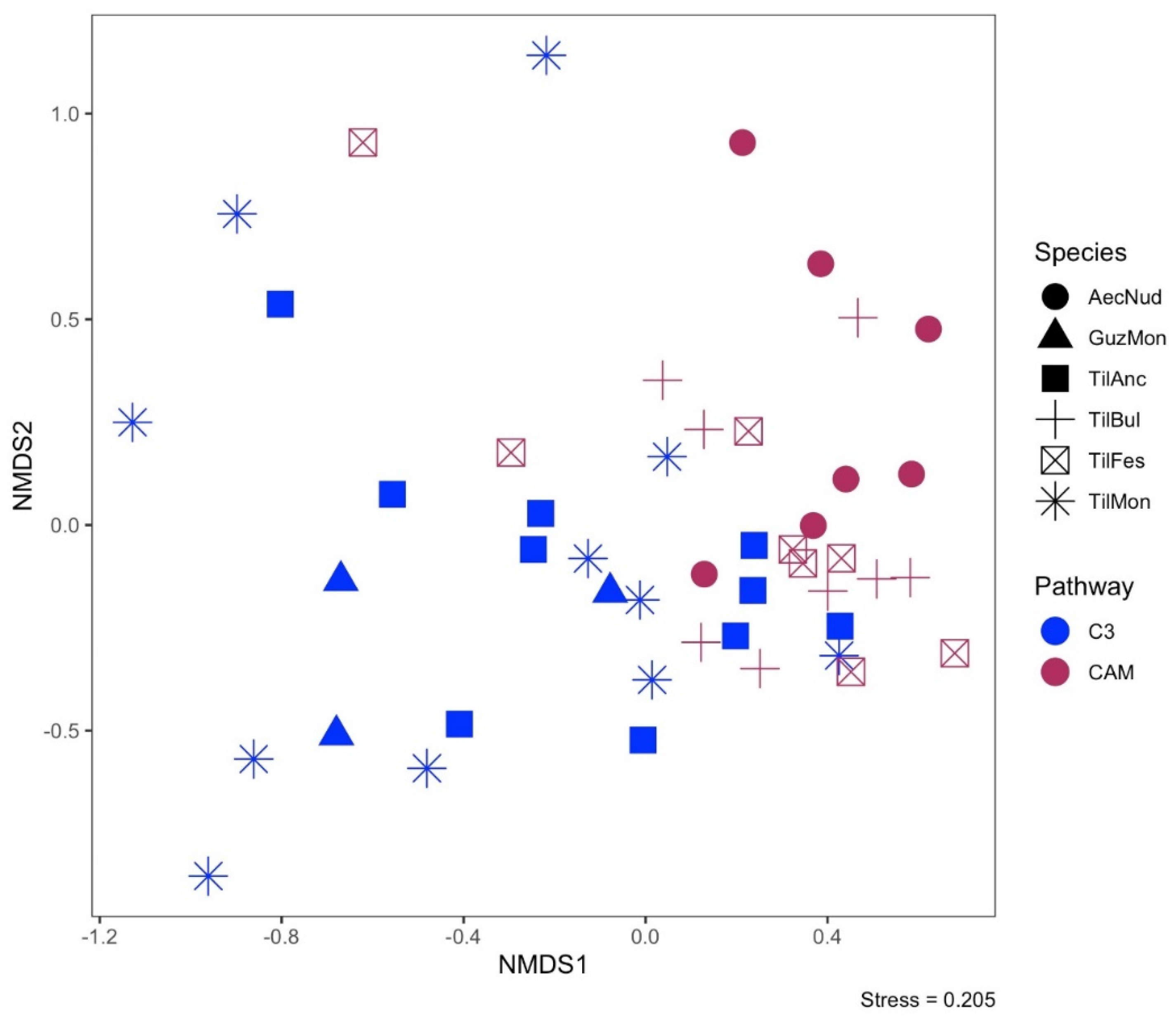
3.2. Endophyte Communities and Host Photosynthetic Pathways
3.3. Endophyte Orders within Bromeliad Species
3.4. Endophyte Communities and Host Ecophysiological Traits
4. Discussion
4.1. CAM vs. C3 Endophyte Communities
4.2. Endophyte Orders in Bromeliad Species
4.3. Leaf Functional Traits and Endophyte Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nieder, J.; Engwald, S.; Barthlott, W. Patterns of neotropical epiphyte diversity. Selbyana 1999, 3, 66–75. [Google Scholar]
- Nadkarni, N.M. Epiphyte biomass and nutrient capital of a neotropical elfin forest. Biotropica 1984, 5, 249–256. [Google Scholar] [CrossRef]
- Zotz, G.; Hietz, P. The physiological ecology of vascular epiphytes: Current knowledge, open questions. J. Exp. Bot. 2001, 52, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Benzing, D.H.; Bennett, B. Bromeliaceae: Profile of an Adaptive Radiation; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Nadkarni, N.M.; Schaefer, D.; Matelson, T.J.; Solano, R. Biomass and nutrient pools of canopy and terrestrial components in a primary and a secondary montane cloud forest, Costa Rica. For. Ecol. Manag. 2004, 198, 223–236. [Google Scholar] [CrossRef]
- Brouard, O.; CÉRÉGhino, R.; Corbara, B.; Leroy, C.; Pelozuelo, L.; Dejean, A.; Carrias, J.-F. Understorey environments influence functional diversity in tank-bromeliad ecosystems. Freshwat. Biol. 2012, 57, 815–823. [Google Scholar] [CrossRef]
- Carrias, J.-F.; Cussac, M.-E.; Corbara, B. A preliminary study of freshwater protozoa in tank bromeliads. J. Trop. Ecol. 2001, 17, 611–617. [Google Scholar] [CrossRef]
- Landell, M.F.; Mautone, J.N.; Valente, P. Biodiversity of yeasts associated to bromeliads in Itapuã park, Viamão/RS. Biociencias 2006, 14, 144–149. [Google Scholar]
- Gonçalves, A.Z.; Hoffmann, F.L.; Mercier, H.; Mazzafera, P.; Romero, G.Q. Phyllosphere bacteria improve animal contribution to plant nutrition. Biotropica 2014, 46, 170–174. [Google Scholar] [CrossRef]
- Torrecillas, E.; Torres, P.; Alguacil, M.; Querejeta, J.; Roldán, A. Influence of habitat and climate variables on arbuscular mycorrhizal fungus community distribution, as revealed by a case study of facultative plant epiphytism under semiarid conditions. Appl. Environ. Microbiol. 2013, 79, 7203–7209. [Google Scholar] [CrossRef] [Green Version]
- Leroy, C.; Carrias, J.-F.; Céréghino, R.; Corbara, B. The contribution of microorganisms and metazoans to mineral nutrition in bromeliads. J. Plant Ecol. 2016, 9, 241–255. [Google Scholar] [CrossRef]
- Lugo, M.A.; Molina, M.G.; Crespo, E.M. Arbuscular mycorrhizas and dark septate endophytes in bromeliads from South American arid environment. Symbiosis 2009, 47, 17–21. [Google Scholar] [CrossRef]
- Lugo, M.A.; Reinhart, K.O.; Menoyo, E.; Crespo, E.M.; Urcelay, C. Plant functional traits and phylogenetic relatedness explain variation in associations with root fungal endophytes in an extreme arid environment. Mycorrhiza 2015, 25, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Unterseher, M.; Gazis, R.; Chaverri, P.; Guarniz, C.F.G.; Tenorio, D.H.Z. Endophytic fungi from Peruvian highland and lowland habitats form distinctive and host plant-specific assemblages. Biodivers. Conserv. 2013, 22, 999–1016. [Google Scholar] [CrossRef]
- Higgins, K.L.; Coley, P.D.; Kursar, T.A.; Arnold, A.E. Culturing and direct PCR suggest prevalent host generalism among diverse fungal endophytes of tropical forest grasses. Mycologia 2011, 103, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Heyduk, K.; Ray, J.N.; Leebens-Mack, J. Leaf anatomy is not correlated to CAM function in a C3+ CAM hybrid species, Yucca gloriosa. Ann. Bot. 2020, 4. [Google Scholar] [CrossRef] [Green Version]
- Del Olmo-Ruiz, M.; Arnold, A.E. Interannual variation and host affiliations of endophytic fungi associated with ferns at La Selva, Costa Rica. Mycologia 2014, 106, 8–21. [Google Scholar] [CrossRef]
- Higgins, K.L.; Arnold, A.E.; Coley, P.D.; Kursar, T.A. Communities of fungal endophytes in tropical forest grasses: Highly diverse host- and habitat generalists characterized by strong spatial structure. Fungal Ecol. 2014, 8, 1–11. [Google Scholar] [CrossRef]
- Donald, J.; Roy, M.; Suescun, U.; Iribar, A.; Manzi, S.; Péllissier, L.; Gaucher, P.; Chave, J. A test of community assembly rules using foliar endophytes from a tropical forest canopy. J. Ecol. 2020, 108, 1605–1616. [Google Scholar] [CrossRef]
- Vincent, J.B.; Weiblen, G.; May, G. Host associations and beta diversity of fungal endophyte communities in New Guinea rainforest trees. Mol. Ecol. 2016, 25, 825–841. [Google Scholar] [CrossRef]
- González-Teuber, M.; Vilo, C.; Guevara-Araya, M.J.; Salgado-Luarte, C.; Gianoli, E. Leaf resistance traits influence endophytic fungi colonization and community composition in a South American temperate rainforest. J. Ecol. 2020, 108, 1019–1029. [Google Scholar] [CrossRef]
- Martin, C.E. Physiological ecology of the Bromeliaceae. Bot. Rev. 1994, 60, 1–82. [Google Scholar] [CrossRef]
- Luttge, U. Ecophysiology of Crassulacean Acid Metabolism (CAM). Ann. Bot. 2004, 93, 629–652. [Google Scholar] [CrossRef] [PubMed]
- Woods, C. Factors influencing the distribution and structure of tropical vascular epiphyte communities at multiple scales. Ph.D. Thesis, Clemson University, Clemson, SC, USA, 2013. [Google Scholar]
- Saunders, M.; Glenn, A.E.; Kohn, L.M. Exploring the evolutionary ecology of fungal endophytes in agricultural systems: Using functional traits to reveal mechanisms in community processes. Evol. Appl. 2010, 3, 525–537. [Google Scholar] [CrossRef]
- Holdridge, L.; Mason, F.; Hatheway, W. Life zone ecology. San José. Trop. Sci. Center 1967, 9, 506. [Google Scholar]
- McDade, L.A.; Bawa, K.S.; Hespenheide, H.A.; Hartshorn, G.S. La Selva: Ecology and Natural History of a Neotropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1994. [Google Scholar]
- Crayn, D.M.; Winter, K.; Schulte, K.; Smith, J.A.C. Photosynthetic pathways in Bromeliaceae: Phylogenetic and ecological significance of CAM and C3 based on carbon isotope ratios for 1893 species. Bot. J. Linn. Soc. 2015, 178, 169–221. [Google Scholar] [CrossRef] [Green Version]
- Woods, C.L.; Cardelús, C.L.; DeWalt, S.J. Microhabitat associations of vascular epiphytes in a wet tropical forest canopy. J. Ecol. 2015, 103, 421–430. [Google Scholar] [CrossRef]
- Cornelissen, J.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.; Reich, P.; Ter Steege, H.; Morgan, H.; Van Der Heijden, M. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A. A method to improve leaf succulence quantification. Braz. Arch. Biol. Technol. 1999, 42. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, N.; Mantuano, D.G.; Mantovani, A. Comparative leaf ecophysiology and anatomy of seedlings, young and adult individuals of the epiphytic aroid Anthurium scandens (Aubl.) Engl. Environ. Exp. Bot. 2010, 68, 314–322. [Google Scholar] [CrossRef]
- Wright, I.; Cannon, K. Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Funct. Ecol. 2001, 15, 351–359. [Google Scholar] [CrossRef] [Green Version]
- Witkowski, E.T.; Lamont, B.B. Leaf specific mass confounds leaf density and thickness. Oecologia 1991, 88, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Sandberg, D.C.; Battista, L.J.; Arnold, A.E. Fungal endophytes of aquatic macrophytes: Diverse host-generalists characterized by tissue preferences and geographic structure. Microb. Ecol. 2014, 67, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef] [PubMed]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Arnold, A.E. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Maddison, W.; Maddison, D. Mesquite: A Modular System for Evolutionary Analysis. Version 2.6, 2.6. Available online: https://www.researchgate.net/publication/235789864_MESQUITE_a_modular_system_for_evolutionary_analysis (accessed on 10 November 2009).
- Ewing, B.; Green, P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome Res. 1998, 8, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; White, J.B.; Miadlikowska, J.; Arnold, A.E.; Miller, M.A.; Kauff, F.; U’Ren, J.M.; May, G.; Lutzoni, F. T-BAS: Tree-Based Alignment Selector toolkit for phylogenetic-based placement, alignment downloads and metadata visualization: An example with the Pezizomycotina tree of life. Bioinformatics 2017, 33, 1160–1168. [Google Scholar] [CrossRef]
- U’ren, J.M.; Dalling, J.W.; Gallery, R.E.; Maddison, D.R.; Davis, E.C.; Gibson, C.M.; Arnold, A.E. Diversity and evolutionary origins of fungi associated with seeds of a neotropical pioneer tree: A case study for analysing fungal environmental samples. Mycol. Res. 2009, 113, 432–449. [Google Scholar] [CrossRef]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.4-0. Available online: https://www.researchgate.net/publication/323265570_vegan_Community_Ecology_Package_R_package_version_24-1_cited_2017_January_25 (accessed on 15 January 2016).
- Martin, C.E.; Wallace, R.S. Photosynthetic pathway variation in leafy members of two subfamilies of the Cactaceae. Int. J. Plant Sci. 2000, 161, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.; Winter, K.; Griffiths, H. The role of CAM in high rainfall cloud forests: an in situ comparison of photosynthetic pathways in Bromeliaceae. Plant Cell Environ. 2002, 25, 1181–1189. [Google Scholar] [CrossRef]
- Inglis, G.D.; Enkerli, J.; Goettel, M.S. Laboratory techniques used for entomopathogenic fungi: Hypocreales. Man. Tech. Invertebr. Pathol. 2012, 2, 18–53. [Google Scholar]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Choong, M.; Lucas, P.; Ong, J.; Pereira, B.; Tan, H.; Turner, I. Leaf fracture toughness and sclerophylly: Their correlations and ecological implications. New Phytol. 1992, 121, 597–610. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.; Falster, D.S.; Garnier, E.; Hikosaka, K.; Lamont, B.B.; Lee, W.; Oleksyn, J.; Osada, N.; et al. Assessing the generality of global leaf trait relationships. New Phytol. 2005, 166, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Read, J.; Sanson, G.; Perez Trautmann, M.F. Leaf traits in Chilean matorral: Sclerophylly within, among, and beyond matorral, and its environmental determinants. Ecol. Evol. 2016, 6, 1430–1446. [Google Scholar] [CrossRef] [Green Version]
- Winkler, M.; Hülber, K.; Mehltreter, K.; Franco, J.G.; Hietz, P. Herbivory in epiphytic bromeliads, orchids and ferns in a Mexican montane forest. J. Trop. Ecol. 2005, 21, 147–154. [Google Scholar] [CrossRef]
- Van Bael, S.; Estrada, C.; Arnold, A.E. Chapter 6: Foliar endophyte communities and leaf traits in tropical trees. In The Fungal Community: Its Organization and Role in the Ecosystem; CRC Press: Boca Raton, FL, USA, 2017; pp. 79–94. [Google Scholar]
- Helander, M.; Ahlholm, J.; Sieber, T.N.; Hinneri, S.; Saikkonen, K. Fragmented environment affects birch leaf endophytes. New Phytol. 2007, 175, 547–553. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Murali, T.S.; Thirunavukkarasu, N.; Govinda Rajulu, M.B.; Venkatesan, G.; Sukumar, R. Endophytic fungal communities in woody perennials of three tropical forest types of the Western Ghats, southern India. Biodivers. Conserv. 2011, 20, 913–928. [Google Scholar] [CrossRef]





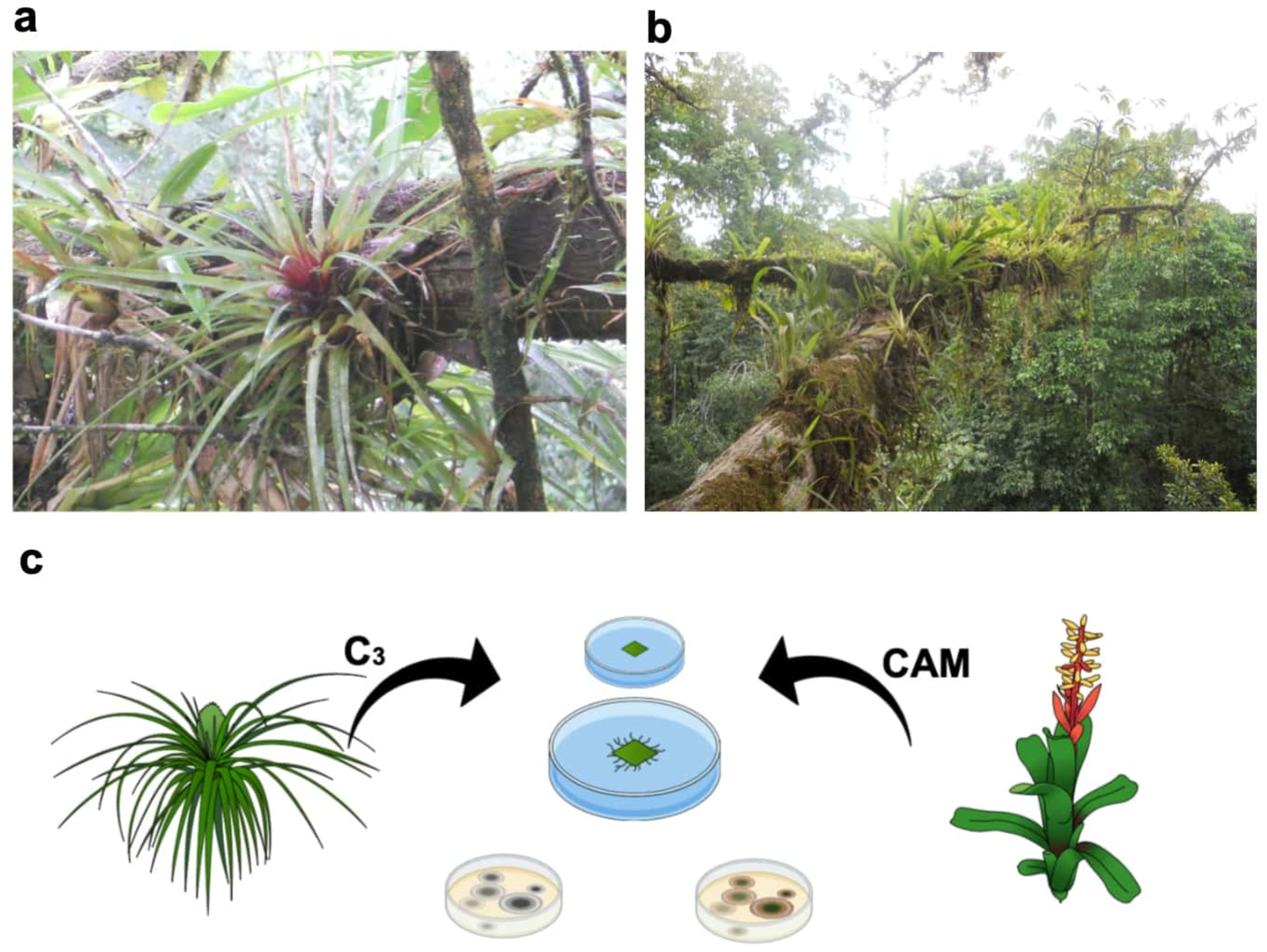{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bromeliads | Endophytes | |||||||
|---|---|---|---|---|---|---|---|---|
| Genus | Species | Code | Photo | Form | N | Abundance | Richness | Diversity |
| Aechmea | A. nudicaulis | AecNud | CAM 1 | Tank | 10 (7) | 0.71 ± 0.18 | 11.3 ± 3.16 | 14.3 ± 6.40 |
| Tillandsia | T. bulbosa | TilBul | CAM 1 | Atmospheric | 10 (8) | 0.78 ± 0.30 | 10.8 ± 4.39 | 11.9 ± 6.39 |
| Tillandsia | T. festucoides | TilFes | CAM 1 | Atmospheric | 9 (8) | 0.78 ± 0.24 | 12.8 ± 3.31 | 17.1 ± 13.5 |
| Guzmania | G. monostachia | GuzMon | C3 1 | Tank | 10 (3) | 0.34 ± 0.24 | 7.00 ± 4.42 | 15.6 ± 3.81 |
| Tillandsia | T. anceps | TilAnc | C3 1 | Tank | 11 (10) | 0.84 ± 0.20 | 11.7 ± 3.63 | 10.9 ± 7.78 |
| Tillandsia | T. monadelpha | TilMon | C3 1 | Tank | 13 (11) | 0.65 ± 0.25 | 11.0 ± 2.75 | 11.7 ± 7.54 |
| Leaf Functional Trait | Formula | Units | Relations to Plant Performance |
|---|---|---|---|
| Specific leaf area (SLA) | Leaf area/dry weight | mm2 mg−1 | Correlates positively with growth rate and negatively with leaf life span 1. |
| Succulence | (Fresh weight-dry weight)/leaf area | g m−2 | Correlates with amount of water storage in plant tissue 2,3. |
| Leaf resistance to fracture | Force/penetrometer circumference | N mm−1 | Indicates carbon investment in structural protection; correlates positively with leaf life span 1 |
| Leaf toughness | (Force/penetrometer circumference)/leaf thickness | N mm−2 | Correlates positively with leaf life span 4,5. |
| Sclerophylly | Dry weight/leaf area | g mm−2 | Correlates positively with leaf life span 5 |
| Rate of Epidermal water loss (EWL) | ∆%Relative water content/h | %RWC h−1 | Relates to cuticle thickness and is low in low water environments 3 |
| Model | Leaf Functional Traits | F | p-Value |
|---|---|---|---|
| Model 1 | Photosynthetic pathway | 4.09 | 0.001 |
| Leaf resistance to fracture | 2.37 | 0.003 | |
| Model 2 | Sclerophylly | 4.430 | 0.001 |
| Leaf resistance to fracture | 2.837 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tellez, P.H.; Woods, C.L.; Formel, S.; Bael, S.A.V. Relationships between Foliar Fungal Endophyte Communities and Ecophysiological Traits of CAM and C3 Epiphytic Bromeliads in a Neotropical Rainforest. Diversity 2020, 12, 378. https://doi.org/10.3390/d12100378
Tellez PH, Woods CL, Formel S, Bael SAV. Relationships between Foliar Fungal Endophyte Communities and Ecophysiological Traits of CAM and C3 Epiphytic Bromeliads in a Neotropical Rainforest. Diversity. 2020; 12(10):378. https://doi.org/10.3390/d12100378
Chicago/Turabian StyleTellez, Peter H., Carrie L. Woods, Stephen Formel, and Sunshine A. Van Bael. 2020. "Relationships between Foliar Fungal Endophyte Communities and Ecophysiological Traits of CAM and C3 Epiphytic Bromeliads in a Neotropical Rainforest" Diversity 12, no. 10: 378. https://doi.org/10.3390/d12100378