
Solanum tuberosum Microtuber Development under Darkness Unveiled through RNAseq Transcriptomic Analysis
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. MTs Transcriptomic-Wide Analysis
2.2. TFs
2.2.1. Up-Regulated TFs
2.2.2. Down-Regulated TFs
2.2.3. PPI Network of Up- and Down-Regulated TFs
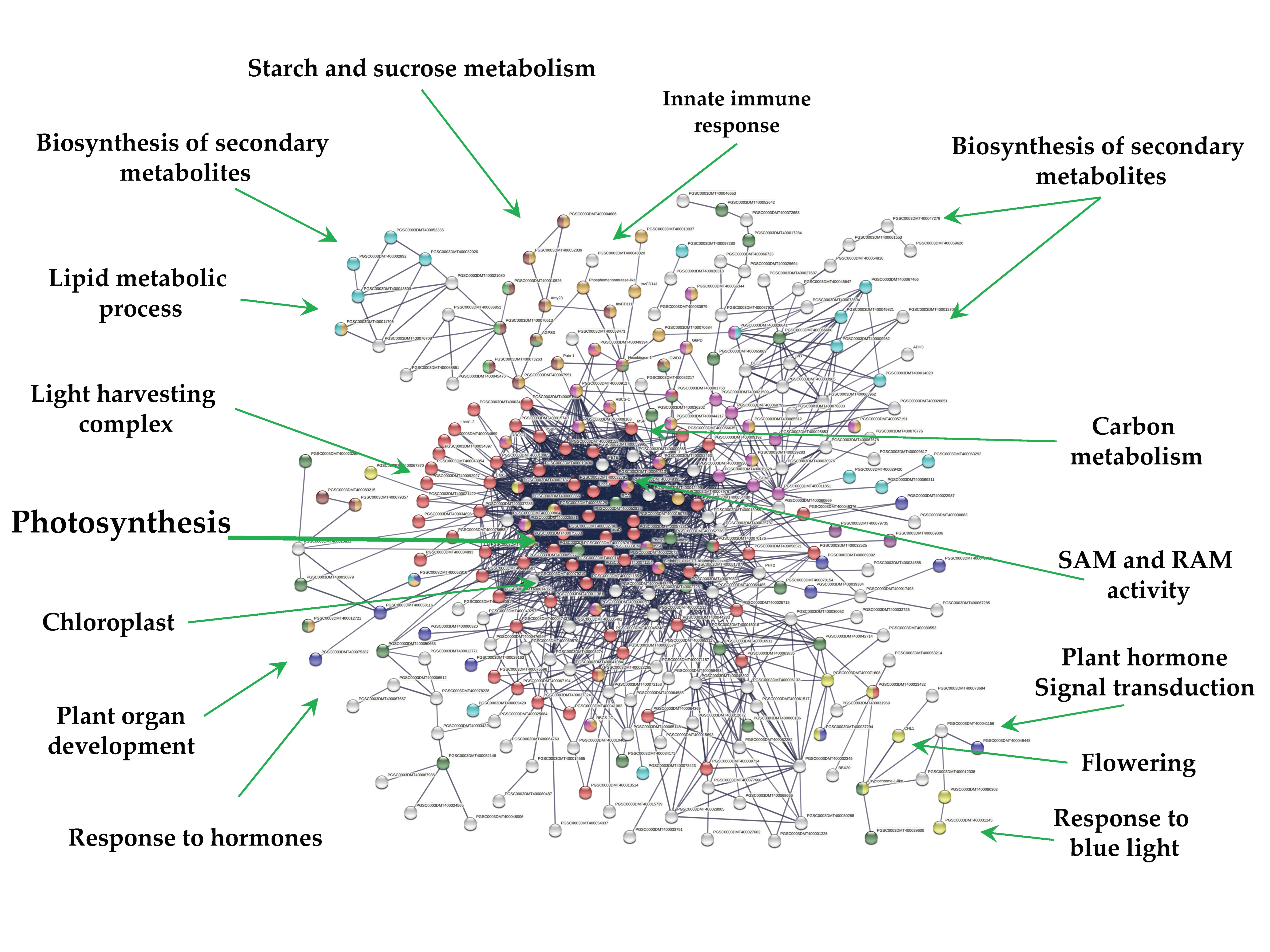
2.3. Up-Regulated Genes
2.4. Down-Regulated Genes
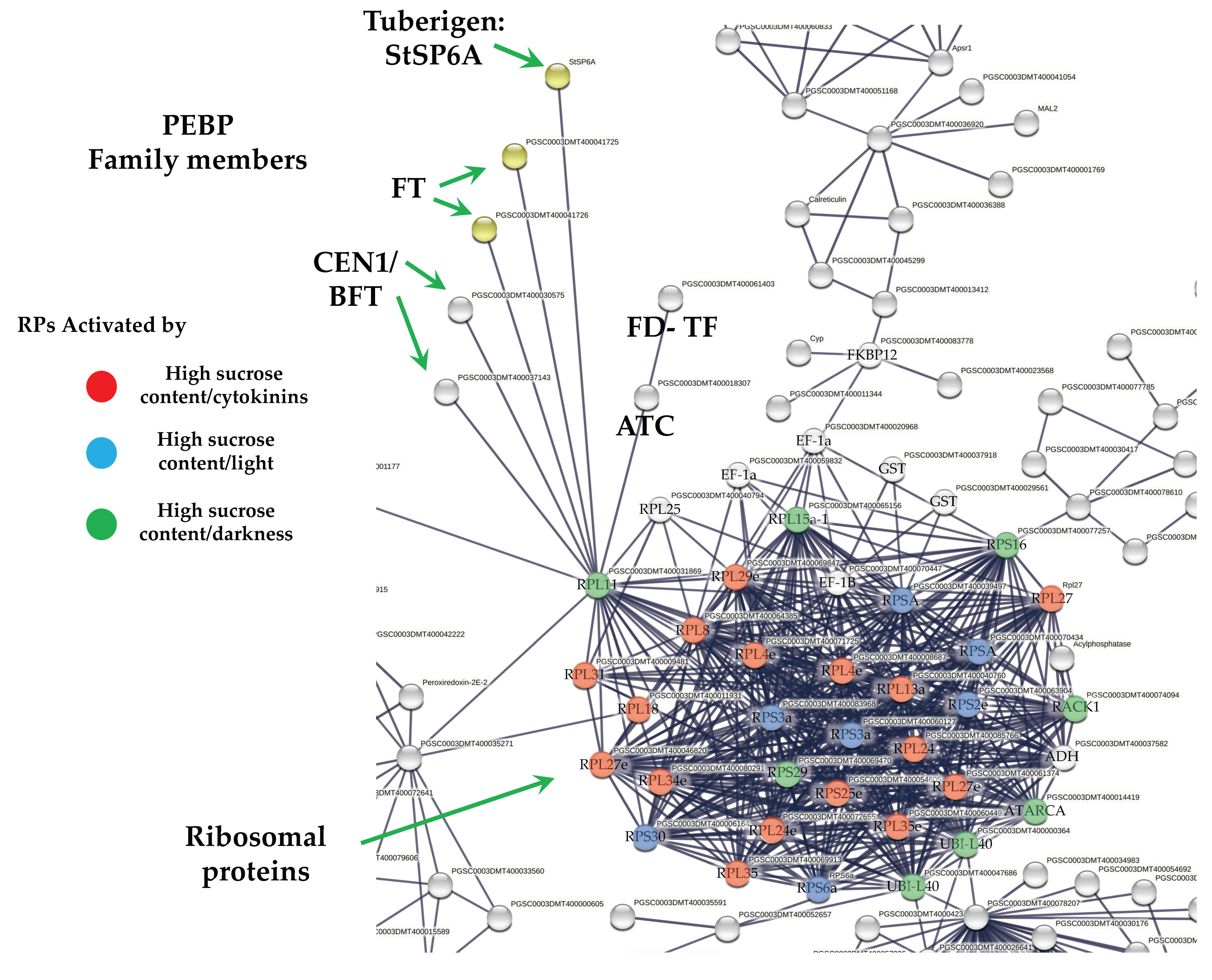
2.5. DEG Related to PEBP Family Members
2.6. DEG Related to RPs
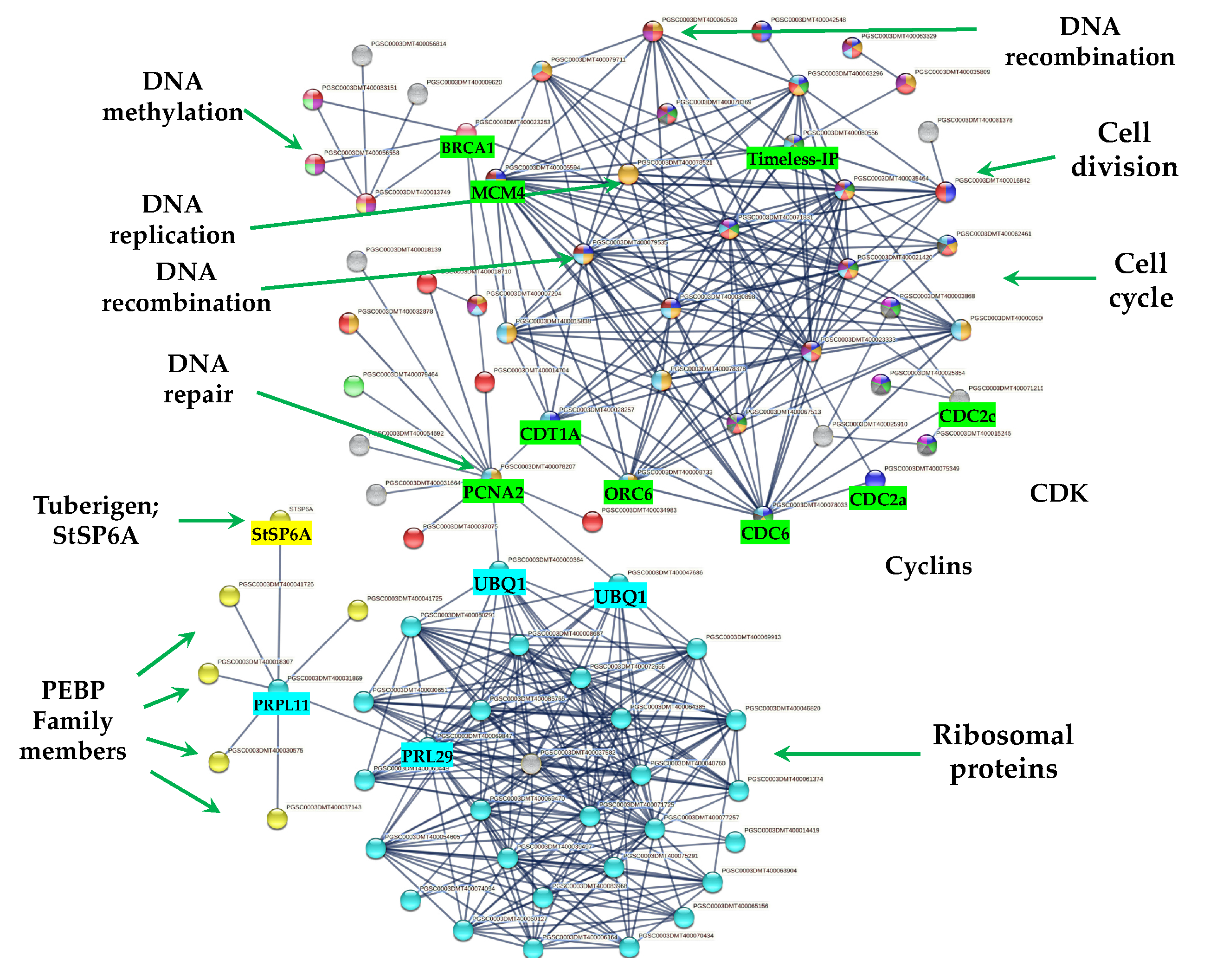
2.7. DEG Related to CC
2.7.1. Cyclins/CDKs
2.7.2. E2F Factors
2.7.3. Histone Binding Proteins
2.7.4. PPI Network Interaction of CC with RPs and Florigens
2.8. DEG Transcripts with Biological Functions
2.9. Validation of the Transcriptome-Wide Analysis
3. Discussion
3.1. PPI Network of Up-Regulated TFs
3.2. PPI Network of Down-Regulated TFs
3.3. Up-Regulated DEG PPI Network
3.3.1. PEBP Family Members and FD TF Interacting with RPs
3.3.2. RPs Cluster
Sucrose Activation of RPs
CK Activation of RPs
Stress Activation of RPs
RPs and Their Relationship with CC
3.3.3. CC Cluster
Cyclins/CDKs
E2F Factors
Histone Binding Proteins
3.4. Validation of Transcriptomic-Wide Analysis by RT-qPCR
4. Materials and Methods
4.1. Plant Material
4.1.1. Potato Shoot Micropropagation
4.1.2. Potato MTs Induction
4.1.3. Transcriptome Sequencing and Assembly
4.1.4. Analysis of DEG
4.1.5. PPI Analysis of Microtuberization
5. Conclusions
- A PPI network of up-regulated TFs revealed that at least six TFs–MYB43, TSF, bZIP27, bZIP43, HAT4, WOX9–may be involved during MTs development.
- Two fundamental biological process essential for life and highly conserved through organisms were found interacting tightly: RPs comprising 29 and CC 117 proteins.
- PEBP members interact with RPs and CC process to activate MTs development under high content of sucrose and gelrite, 2iP under darkness.
- Further experiments by yeast two-hybrid screening approach of genome edited up- and down-regulated PEBP members with RPL11 are required to demonstrate our model of MTs development under darkness.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization Corporate Statistical Database. FAOSTAT. 2022. Available online: https://www.fao.org/faostat/es/#data/QCL/visualize (accessed on 26 July 2022).
- Khosa, J.; Bellinazzo, F.; Kamenetsky, R.; Macknight, R.; Immink, R.G.H. PHOSPHATIDYLETHANOLAMONE-BINDING PROTEINS: The conductors of dual reproduction in plants with vegetative storage organs. J. Exp. Bot. 2021, 72, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Dutt, S.; Manjul, A.S.; Raigond, P.; Singh, B.; Siddappa, S.; Bhardwaj, V.; Kawar, P.G.; Patil, V.U.; Kardile, H.B. Key Players Associated with Tuberization in Potato: Potential Candidates for Genetic Engineering. Crit. Rev. Biotechnol. 2017, 37, 942–957. [Google Scholar] [CrossRef] [PubMed]
- Kondhare, K.R.; Malankar, N.N.; Devani, R.S.; Banerjee, A.K. Genome-Wide Transcriptome Analysis Reveals Small RNA Profiles Involved in Early Stages of Stolon-to-Tuber Transitions in Potato under Photoperiodic Conditions. BMC Plant Biol. 2018, 18, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zierer, W.; Rüscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and Tuberous Root Development. Annu. Rev. Plant Biol. 2021, 72, 551–580. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Coleman, W.K.; Coleman, S.E. Potato Microtuber Production and Performance: A Review. Am. J. Pot. Res. 2003, 80, 103–115. [Google Scholar] [CrossRef]
- Hannapel, D.J. Signalling the Induction of Tuber Formation. In Potato Biology and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2007; pp. 237–256. [Google Scholar] [CrossRef]
- Vinterhalter, D.; Dragicevic, I.; Vinterhalter, B. Potato in vitro culture techniques and biotechnology. Fruit Veg. Cereal Sci. Biotechnol. (Spec. Issue 1) 2008, 2, 16–45. [Google Scholar]
- Coleman, W.K.; Donnelly, D.J.; Coleman, S.E. Potato Microtubers as Research Tools: A Review. Am. J. Pot. Res. 2001, 78, 47–55. [Google Scholar] [CrossRef]
- Herrera-Isidron, L.; Valencia-Lozano, E.; Rosiles-Loeza, P.Y.; Robles-Hernández, M.G.; Napsuciale-Heredia, A.; Cabrera-Ponce, J.L. Gene Expression Analysis of Microtubers of Potato Solanum Tuberosum L. Induced in Cytokinin Containing Medium and Osmotic Stress. Plants 2021, 10, 876. [Google Scholar] [CrossRef]
- Crookshanks, M.; Emmersen, J.; Welinder, K.G.; Lehmann-Nielsen, K. The potato tuber transcriptome analysis of 6067 expressed sequence tags. FEBS Lett. 2001, 506, 123–126. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Song, W.; Zhou, J.; Wang, X.; Xie, C.; Gao, X.; Xie, T.; Liu, J. Transcriptome analysis reveals novel genes potentially involved in photoperiodic tuberization in potato. Genomics 2013, 102, 388–396. [Google Scholar] [CrossRef]
- Kumar, J.; Sapna, T.; Sundaresha, D.S.; Chandel, P.; Ali, N.; Singh, B.; Bhardwaj, V.; Singh, B.P. Microarray analysis of gene expression patterns in the leaf during potato tuberization in the potato somatic hybrid Solanum tuberosum and Solanum etuberosum. Genome 2014, 58, 305–313. [Google Scholar] [CrossRef]
- Gong, L.; Zhang, H.; Gan, X.; Zhang, L.; Chen, Y.; Nie, F.; Shi, L.; Li, M.; Guo, Z.; Zhang, G.; et al. Transcriptome Profiling of the Potato (Solanum tuberosum L.) Plant under Drought Stress and Water-Stimulus Conditions. PLoS ONE 2015, 10, e0128041. [Google Scholar] [CrossRef]
- Sharma, P.; Lin, T.; Hannapel, D.J. Targets of the StBEL5 Transcription Factor Include the FT Ortholog StSP6A. Plant Physiol. 2016, 170, 310–324. [Google Scholar] [CrossRef] [Green Version]
- Vulavala, V.K.R.; Fogelman, E.; Faigenboim, A.; Shoseyov, O.; Ginzberg, I. The Transcriptome of Potato Tuber Phellogen Reveals Cellular Functions of Cork Cambium and Genes Involved in Periderm Formation and Maturation. Sci. Rep. 2019, 9, 10216. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.N.; Doudna Cate, J.H. The Structure and Function of the Eukaryotic Ribosome. Cold Spring Harb. Perspect. Biol. 2012, 4, a011536. [Google Scholar] [CrossRef]
- Inzé, D.; De-Veylder, L. Cell Cycle Regulation in Plant Development. Annu. Rev. Genet. 2006, 40, 77–105. [Google Scholar] [CrossRef]
- Timsit, Y.; Sergeant-Perthuis, G.; Bennequin, D. Evolution of ribosomal protein network architectures. Sci. Rep. 2021, 11, 625. [Google Scholar] [CrossRef]
- Gamm, M.; Peviani, A.; Honsel, A.; Snel, B.; Smeekens, S.; Hanson, J. Increased Sucrose Levels Mediate Selective mRNA Translation in Arabidopsis. BMC Plant Biol. 2014, 14, 306. [Google Scholar] [CrossRef] [Green Version]
- Brenner, W.G.; Schmülling, T. Transcript Profiling of Cytokinin Action in Arabidopsis Roots and Shoots Dcovers Largely Similar but Also Organ-Specific Responses. BMC Plant Biol. 2012, 12, 112. [Google Scholar] [CrossRef] [Green Version]
- Latchman, D.S. Transcription factors: An overview. Int. J. Biochem. Cell Biol. 1997, 29, 1305–1312. [Google Scholar] [CrossRef]
- Nicolas, M.; Torres-Pérez, R.; Wahl, V.; Cruz-Oró, E.; Rodríguez-Buey, M.L.; Zamarreño, A.M.; Martín-Jouve, B.; García-Mina, J.M.; Oliveros, J.C.; Prat, S.; et al. Spatial control of potato tuberization by the TCP transcription factor BRANCHED1b. Nat. Plants 2022, 8, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Jia, Y.; Zhang, J.; Li, H.; Cheng, L.; Wang, P.; Bao, Z.; Liu, Z.; Feng, S.; Zhu, X.; et al. Genome evolution and diversity of wild and cultivated potatoes. Nature 2022, 606, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Lai, T.H.; Kim, D.R. A functional Network Model of the Metastasis Supressor PEBP1/RKIP and its Regulators in Breast Cancer Cells. Cancers 2021, 13, 6098. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.; Seitz, T.; Li, S.; Janosch, P.; McFerran, B.; Kaiser, C.; Fee, F.; Katsanakis, K.D.; Rose, D.W.; Mischak, H.; et al. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature 1999, 401, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Al-Mulla, F.; Bitar, M.S.; Taqi, Z.; Rath, O.; Kolch, W. RAF kinase activity signalling by (RKIP) modulates cell cycle kinetics and motility. Mol. Biosyst. 2011, 7, 928–941. [Google Scholar] [CrossRef]
- Voogd, C.; Brian, L.A.; Wang, T.; Allan, A.C.; Varkony-Gasic, E. Three FT and multiple CEN and BFT genes regulate maturity, flowering, and vegetative phenology in kiwifruit. J. Exp. Bot. 2017, 68, 1539–1553. [Google Scholar] [CrossRef] [Green Version]
- Navarro, C.; Abelenda, J.A.; Cruz-Oró, E.; Cuéllar, C.A.; Tamaki, S.; Silva, J.; Shimamoto, K.; Prat, S. Control of Flowering and Storage Organ Formation in Potato by FLOWERING LOCUS T. Nature 2011, 478, 119–122. [Google Scholar] [CrossRef]
- Moraes, T.S.; Dornelas, M.C.; Martinelli, A.P. FT/TFL1: Calibrating Plant Architecture. Front. Plant Sci. 2019, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Collani, S.; Neumann, M.; Yant, L.; Schmid, M. FT Modulates Genome-wide DNA-Binding of the bZIP Transcription Factor FD. Plant Physiol. 2019, 180, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Purwestri, Y.A.; Susanto, F.A.; Tsuji, H. Hd3a florigen recruits different proteins to reveal its function in plant growth and development. Intech 2017, 4, 49–67. [Google Scholar] [CrossRef]
- Wang, E.; Liu, T.; Sun, X.; Jing, S.; Zhou, T.; Liu, T.; Song, B. Profiling of the Candidate Interacting Proteins of SELF-PRUNING 6A (SP6A) in Solanum tuberosum. Int J. Mol. Sci. 2022, 15, 9126. [Google Scholar] [CrossRef] [PubMed]
- Abelenda, J.A.; Bergonzi, S.; Oortwijn, M.; Sonnewald, S.; Du, M.; Visser, R.G.; Bachem, C.W. Source-sink regulation is mediated by interaction of an FT homolog with a SWEET protein in potato. Curr. Biol. 2019, 29, 1178–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plantenga, F.D.M.; Bergonzi, S.; Abelenda, J.A.; Bachem, C.W.B.; Visser, R.G.F.; Heuvelink, E.; Marcelis, L.F.M. The tuberization signal StSP6A represses flower bud development in potato. J. Exp. Bot. 2019, 70, 937–948. [Google Scholar] [CrossRef]
- Navarro, C.; Cruz-Oró, E.; Pratt, S. Conserved function of flowering (FT) homologues as signals for storage organ differentiation. Curr. Opin. Plant Biol. 2015, 23, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Hastilestari, B.R.; Lorenz, J.; Reid, S.; Hofman, J.; Pscheidt, D.; Sonnewald, U.; Sonnewald, S. Deciphering source and sink responses of potato plants (Solanum tuberosum L.) to elevated temperatures. Plant Cell Envirom. 2018, 41, 2600–2616. [Google Scholar] [CrossRef] [PubMed]
- Lehretz, G.G.; Sonnewald, S.; Hornyik, C.; Corral, J.M.; Sonnewald, U. Post-transcriptional Regulation of FLOWERING LOCUS T Modulates Heat-Dependent Source-Sink Development in Potato. Curr. Biol. 2019, 29, 1614–1624.e3. [Google Scholar] [CrossRef] [PubMed]
- Salvato, F.; Havelund, J.F.; Chen, M.; Rao, R.S.P.; Rogowska-Wrzesinska, A.; Jensen, O.N.; Gang, D.R.; Thelen, J.J.; Møller, I.M. The Potato Tuber Mitochondrial Proteome. Plant Physiol. 2014, 16, 637–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, J.P.; Cankar, K.; Scheffer, S.J.; Beenen, H.G.; Shepherd, L.V.T.; Stewart, D.; Davies, H.V.; Wilkockson, S.J.; Leifert, C.; Gruden, K.; et al. Transcriptome Analysis of Potato Tubers Effects of Different Agricultural Practices. J. Agric. Food Chem. 2009, 57, 1612–1623. [Google Scholar] [CrossRef] [PubMed]
- Jing, S.; Sun, X.; Yu, L.; Wang, E.; Cheng, Z.; Liu, H.; Jiang, P.; Qin, J.; Begum, S.; Song, B. Transcription factor StABI5-like 1 binding to the FLOWERING LOCUS T homologs promotes early maturity in potato. Plant Physiol. 2022, 18, 1677–1693. [Google Scholar] [CrossRef]
- Mhawech, P. 14-3-3 proteins-an update. Cell Res. 2005, 15, 228–236. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, W.; Yu, H.; Peng, J.; Hu, Z.; Chen, L. The role of 14-3-3 proteins in plant growth and response to abiotic stress. Plant Cell Rep. 2022, 41, 833–852. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; van Esse, W.; Mulki, M.A.; Kirschner, G.; Zhong, J.; Simon, R.; von Korff, M. CENTRORADIALIS Interacts with FLOWERING LOCUS T-Like Genes to Control Floret Development and Grain Number. Plant Physiol. 2019, 180, 1013–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Campbell, R.; Ducreux, L.J.M.; Morris, J.; Hedley, P.E.; Mellado-Ortega, E.; Roberts, A.G.; Stephens, J.; Bryan, G.J.; Torrance, L.; et al. TERMINAL FLOWER-1/CENTRORADIALIS inhibits tuberization via protein interaction with the tuberigen activation complex. Plant J. 2020, 103, 2263–2278. [Google Scholar] [CrossRef] [PubMed]
- Morris, W.L.; Alamar, M.C.; Lopez-Cobollo, R.M.; Castillo Cañete, J.; Bennett, M.; Van der Kaay, J.; Stevens, J.; Kumar Sharma, S.; McLean, K.; Thompson, A.J.; et al. A member of the TERMINAL FLOWER 1/CENTRORADIALIS gene family controls sprout growth in potato tubers. J. Exp. Bot. 2019, 70, 835–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abelenda, J.A.; Cruz-Oró, E.; Franco-Zorrilla, J.M.; Prat, S. Potato StCONSTANS-Like1 Suppresses Storage Organ Formation by Directly Activating the FT-like StSP5G Repressor. Curr. Biol. 2016, 26, 872–881. [Google Scholar] [CrossRef] [Green Version]
- Teo, C.J.; Takahashi, K.; Shimizu, K.; Shimamoto, K.; Taoka, K.I. Potato Tuber Induction is Regulated by Interactions Between Components of a Tuberigen Complex. Plant Cell Physiol. 2017, 58, 365–374. [Google Scholar] [CrossRef]
- Taylor, M.A.; Arif, S.A.M.; Pearce, S.R.; Davies, H.V.; Kumar, A.; George, L.A. Differential Expression and Sequence Analysis of Ribosomal Protein Genes Induced in Stolon Tips of Potato (Solanum Tuberosum L.) during the Early Stages of Tuberization. Plant Physiol. 1992, 100, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Van Lijsebettens, M.; Vanderhaeghen, R.; De Block, M.; Bauw, G.; Villarroel, R.; Van Montagu, M. An S18 Ribosomal Protein Gene Copy at the Arabidopsis PFL Locus Affects Plant Development by Its Specific Expression in Meristems. EMBO J. 1994, 13, 3378–3388. [Google Scholar] [CrossRef]
- Tsugeki, R.; Kochieva, E.Z.; Fedoroff, N.V. A Transposon Insertion in the Arabidopsis SSR16 Gene Causes an Embryo-Defective Lethal Mutation. Plant J. 1996, 10, 479–489. [Google Scholar] [CrossRef]
- Zinn, A.R.; Ross, J.L. Turner Syndrome and Haploinsufficiency. Curr. Opin. Genet. Dev. 1998, 8, 322–327. [Google Scholar] [CrossRef]
- Lambertsson, A. The Minute Genes in Drosophila and Their Molecular Functions. Adv. Genet. 1998, 38, 69–134. [Google Scholar] [CrossRef] [PubMed]
- Volarević, S.; Stewart, M.J.; Ledermann, B.; Zilberman, F.; Terracciano, L.; Montini, E.; Grompe, M.; Kozma, S.C.; Thomas, G. Proliferation, But Not Growth, Blocked by Conditional Deletion of 40 S Ribosomal Protein S6. Science 2000, 288, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Kim, G.-T.; Shinozaki, K. Disruption of an Arabidopsis Cytoplasmic Ribosomal Protein S13-Homologous Gene by Transposon-Mediated Mutagenesis Causes Aberrant Growth and Development. Plant J. 2000, 22, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Seidel, F.; Beine-Golovchuk, O.; Hsieh, Y.-C.; Kopka, J. Systematic Review of Plant Ribosome Heterogeneity and Specialization. Front. Plant Sci. 2020, 11, 948. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Pezza, A.; Biarc, J.; Burlingame, A.L.; Casati, P. Plant L10 Ribosomal Proteins Have Different Roles during Development and Translation under Ultraviolet-B Stress. Plant Physiol. 2010, 153, 1878–1894. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lan, P.; Gao, H.; Zheng, L.; Li, W.; Schmidt, W. Expression Changes of Ribosomal Proteins in Phosphate- and Iron-Deficient Arabidopsis Roots Predict Stress-Specific Alterations in Ribosome Composition. BMC Genom. 2013, 14, 783. [Google Scholar] [CrossRef] [Green Version]
- Cheong, B.E.; Beine-Golovchuk, O.; Gorka, M.; Ho, W.W.H.; Martinez-Seidel, F.; Firmino, A.A.P.; Skirycz, A.; Roessner, U.; Kopka, J. Arabidopsis REI-LIKE Proteins Activate Ribosome Biogenesis during Cold Acclimation. Sci. Rep. 2021, 11, 2410. [Google Scholar] [CrossRef]
- Zhang, C.; Li, H.; Yuan, C.; Liu, S.; Li, M.; Zhu, J.; Lin, X.; Lu, Y.; Guo, X. CKB 1 Regulates Expression of Ribosomal Protein L10 Family Gene and Plays a Role in UV -B Response. Plant Biol. J. 2020, 22, 143–152. [Google Scholar] [CrossRef]
- Moin, M.; Bakshi, A.; Saha, A.; Dutta, M.; Madhav, S.M.; Kirti, P.B. Rice Ribosomal Protein Large Subunit Genes and Their Spatio-Temporal and Stress Regulation. Front. Plant Sci. 2016, 7, 1284. [Google Scholar] [CrossRef] [Green Version]
- Moin, M.; Bakshi, A.; Madhav, M.S.; Kirti, P.B. Expression Profiling of Ribosomal Protein Gene Family in Dehydration Stress Responses and Characterization of Transgenic Rice Plants Overexpressing RPL23A for Water-Use Efficiency and Tolerance to Drought and Salt Stresses. Front. Chem. 2017, 5, 97. [Google Scholar] [CrossRef]
- Moin, M.; Saha, A.; Bakshi, A.; Divya, D.; Madhav, M.S.; Kirti, P.B. Study on Transcriptional Responses and Identification of Ribosomal Protein Genes for Potential Resistance against Brown Planthopper and Gall Midge Pests in Rice. Curr. Genom. 2021, 22, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Gonzali, S.; Loreti, E.; Solfanelli, C.; Novi, G.; Alpi, A.; Perata, P. Identification of Sugar-Modulated Genes and Evidence for in Vivo Sugar Sensing in Arabidopsis. J. Plant Res. 2006, 119, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Baena-González, E.; Rolland, F.; Thevelein, J.M.; Sheen, J. A Central Integrator of Transcription Networks in Plant Stress and Energy Signalling. Nature 2007, 44, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Retzlaff, K.; Höhne, M.; Günther, M.; Stitt, M. Global Transcript Levels Respond to Small Changes of the Carbon Status during Progressive Exhaustion of Carbohydrates in Arabidopsis Rosettes. Plant Physiol. 2008, 146, 1834–1861. [Google Scholar] [CrossRef] [Green Version]
- Hummel, M.; Cordewener, J.H.G.; de Groot, J.C.M.; Smeekens, S.; America, A.H.P.; Hanson, J. Dynamic Protein Composition of Arabidopsis Thaliana Cytosolic Ribosomes in Response to Sucrose Feeding as Revealed by Label Free MSE Proteomics. Proteomics 2012, 12, 1024–1038. [Google Scholar] [CrossRef]
- Berridge, M.V.; Ralph, R.K.; Letham, D.S. The Binding of Kinetin to Plant Ribosomes. Biochem. J. 1970, 119, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.E.; Erion, J.L.A. Cytokinin Binding Protein from Higher Plant Ribosomes. Biochem. Biophys. Res. Commun. 1975, 64, 694–700. [Google Scholar] [CrossRef]
- Tepfer, D.A.; Fosket, D.E. Hormone-Mediated Translational Control of Protein Synthesis in Cultured Cells of Glycine Max. Dev. Biol. 1978, 62, 486–497. [Google Scholar] [CrossRef]
- Feierabend, J.; de Boer, J. Comparative Analysis of the Action of Cytokinin and Light on the Formation of Ribulosebisphosphate Carboxylase and Plastid Biogenesis. Planta 1978, 142, 75–82. [Google Scholar] [CrossRef]
- Yakovleva, L.; Kulaeva, O. The effect of phytohormones on phosphorylation of ribosomal proteins in detached pumpkin cotyledons. Biochem. Physiol. Pflanz. 1987, 182, 359–365. [Google Scholar] [CrossRef]
- Cherepneva, G.N.; Schmidt, K.-H.; Kulaeva, O.N.; Oelmüller, R.; Kusnetsov, V.V. Expression of the Ribosomal Proteins S14, S16, L13a and L30 Is Regulated by Cytokinin and Abscisic Acid. Plant Sci. 2003, 165, 925–932. [Google Scholar] [CrossRef]
- Karunadasa, S.S.; Kurepa, J.; Shull, T.E.; Smalle, J.A. Cytokinin-induced Protein Synthesis Suppresses Growth and Osmotic Stress Tolerance. New Phytol. 2020, 227, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Hulm, J.L.; McIntosh, K.B.; Bonham-Smith, P.C. Variation in Transcript Abundance among the Four Members of the Arabidopsis thaliana RIBOSOMAL PROTEIN S15a Gene Family. Plant Science 2005, 169, 267–278. [Google Scholar] [CrossRef]
- Chotewutmontri, P.; Stiffler, N.; Watkins, K.P.; Barkan, A. Ribosome Profiling in Maize. In Maize; Methods in Molecular, Biology; Lagrimini, L.M., Ed.; Springer: New York, NY, USA, 2018; Volume 1676, pp. 165–183. [Google Scholar] [CrossRef]
- Lei, L.; Shi, J.; Chen, J.; Zhang, M.; Sun, S.; Xie, S.; Li, X.; Zeng, B.; Peng, L.; Hauck, A.; et al. Ribosome Profiling Reveals Dynamic Translational Landscape in Maize Seedlings under Drought Stress. Plant J. 2015, 84, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Lei, L.; Shi, J.; Wang, X.; Chen, J.; Xue, M.; Sun, S.; Zhan, B.; Xia, Z.; Jiang, N.; et al. Characterization of Maize Translational Responses to Sugarcane Mosaic Virus Infection. Virus Res. 2019, 259, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Shiraku, M.L.; Magwanga, R.O.; Cai, X.; Kirungu, J.N.; Xu, Y.; Mehari, T.G.; Hou, Y.; Wang, Y.; Agong, S.G.; Peng, R.; et al. Functional Characterization of GhACX3 Gene Reveals Its Significant Role in Enhancing Drought and Salt Stress Tolerance in Cotton. Front. Plant Sci. 2021, 12, 658755. [Google Scholar] [CrossRef]
- Mehdi, R.; Mojtaba, K.; Mojtaba, M.; Sanam, S.C. Identification of Drought-Responsive Proteins of Sensitive and Tolerant Tea (Camellia sinensis L.) Clones under Normal and Drought Stress Conditions. Curr. Prot. 2020, 17, 227–240. [Google Scholar] [CrossRef]
- Thapa, M.; Bommakanti, A.; Shamsuzzaman, M.; Gregory, B.; Samsel, L.; Zengel, J.M.; Lindahl, L. Repressed Synthesis of Ribosomal Proteins Generates Protein-Specific Cell Cycle and Morphological Phenotypes. MBoC. 2013, 24, 3620–3633. [Google Scholar] [CrossRef] [Green Version]
- Maitra, N.; He, C.; Blank, H.M.; Tsuchiya, M.; Schilling, B.; Kaeberlein, M.; Aramayo, R.; Kennedy, B.K.; Polymenis, M. Translational Control of One-Carbon Metabolism Underpins Ribosomal Protein Phenotypes in Cell Division and Longevity. eLife 2020, 9, e53127. [Google Scholar] [CrossRef]
- Piper, M.D.; Hong, S.-P.; Ball, G.E.; Dawes, I.W. Regulation of the Balance of One-Carbon Metabolism in Saccharomyces Cerevisiae. J. Biol. Chem. 2000, 275, 30987–30995. [Google Scholar] [CrossRef] [Green Version]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Cortijo, S.; Korsbo, N.; Roszak, P.; Schiessl, K.; Gurzadyan, A.; Wightman, R.; Jönsson, H.; Meyerowitz, E. Molecular mechanism of cytokinin-activated cell division in Arabidopsis. Science 2021, 26, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.; Hwang, K.; Cho, S.; Lee, M.; Kil, E.J.; Choi, S.; Hahn, B.S.; Kim, D.; Auh, C.K.; Lee, S. Expression analysis of D-type cyclin in potato (Solanum tuberosum L.) under different culture conditions. Acta Physiol. Plant. 2016, 38, 36. [Google Scholar] [CrossRef]
- Lara-Gonzalez, P.; Kim, T.; Oegema, K.; Corbett, K.; Desai, A. A tripartite mechanism catalyzes Mad2-Cdc20 assembly at unattached kinetochores. Science 2021, 374, 64–67. [Google Scholar] [CrossRef]
- Leu, W.M.; Cao, X.L.; Wilson, T.J.; Snustad, D.P.; Chua, N.H. Phytochrome A and phytochrome B mediate the hypocotyl-specific down-regulation of TUB1 by light in arabidopsis. Plant Cell 1995, 7, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Demidov, D.; Van Damme, D.; Geelen, D.; Blattner, F.R.; Houben, A. Identification and dynamics of two classes of aurora-like kinases in Arabidopsis and other plants. Plant Cell 2005, 17, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Cai, G.; Cresti, M. Are kinesins required for organelle trafficking in plant cells? Front. in Plant Sci. 2012, 3, 170. [Google Scholar] [CrossRef] [Green Version]
- Weaver, R.L.; Limzerwala, J.F.; Naylor, R.M.; Jeganathan, K.B.; Baker, D.J.; van Deursen, J.M. BubR1 alterations that reinforce mitotic surveillance act against aneuploidy and cancer. Elife 2016, 5, e16620. [Google Scholar] [CrossRef]
- Hemerly, A.; Engler, J.D.A.; Bergounioux, C.; Van Montagu, M.; Engler, G.; Inzé, D.; Ferreira, P. Dominant negative mutants of the Cdc2 kinase uncouple cell division from iterative plant development. EMBO J. 1995, 14, 3925–3936. [Google Scholar] [CrossRef]
- Boruc, J.; Van den Daele, H.; Hollunder, J.; Rombauts, S.; Mylle, E.; Hilson, P.; Russinova, E. Functional modules in the Arabidopsis core cell cycle binary protein–protein interaction network. Plant Cell 2010, 22, 1264–1280. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, P.; Stevens, R.; Perennes, C.; Jasinski, S.; Cella, R.; Tremousaygue, D.; Bergounioux, C. AtE2F-a and AtDP-a, members of the E2F family of transcription factors, induce Arabidopsis leaf cells to re-enter S phase. Mol. Genet. Genom. 2002, 266, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Z.; Tsai, S.Y.; Leone, G. Emerging roles of E2Fs in cancer: An exit from cell cycle control. Nat. Rev. Cancer 2009, 11, 785–797. [Google Scholar] [CrossRef] [Green Version]
- Schulman, I.; Bloom, K.S. Centromeres: An integrated protein/DNA complex required for chromosome movement. Annu. Rev. of Cell Biol. 1991, 7, 311–336. [Google Scholar] [CrossRef] [PubMed]
- Pinto, I.; Winston, F. Histone H2A is required for normal centromere function in Saccharomyces cerevisiae. EMBO J. 2002, 19, 1598–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios-Doria, J.; Velkova, A.; Dapic, V.; Galán-Caridad, J.M.; Dapic, V.; Carvalho, M.A.; Melendez, J.; Monteiro, A.N.A. Ectopic expression of histone H2AX mutants reveals a role for its post-translational modifications. Cancer Biol. Ther. 2009, 8, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Von Mering, C. STRING v10: Protein–protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Bombarely, A.; Menda, N.; Tecle, I.Y.; Buels, R.M.; Strickler, S.; Fischer-York, T.; Mueller, L.A. The Sol Genomics Network (solgenomics. net): Growing tomatoes using Perl. Nucleic Acids Res. 2010, 39, D1149–D1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef] [PubMed]
- Sherry, S.T.; Ward, M.H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, G.-H.; Liu, M.-J.; Xiong, Y.; Sheen, J.; Wu, S.-H. TOR and RPS6 Transmit Light Signals to Enhance Protein Translation in Deetiolating Arabidopsis Seedlings. Proc. Natl. Acad. Sci. USA 2018, 115, 12823–12828. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-K.; Kim, S.; Shin, Y.; Hur, Y.-S.; Kim, W.-Y.; Lee, M.-S.; Cheon, C.-I.; Verma, D.P.S. Ribosomal Protein S6, a Target of Rapamycin, Is Involved in the Regulation of RRNA Genes by Possible Epigenetic Changes in Arabidopsis. J. Biol. Chem. 2014, 289, 3901–3912. [Google Scholar] [CrossRef] [Green Version]
- Tzafrir, I. The Arabidopsis SeedGenes Project. Nucleic Acids Res. 2003, 31, 90–93. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, J.; Jia, M.; Xu, N.; Liang, S.; Shao, J.; Qi, Y.; Liu, X.; An, L.; Yu, F. Balance between Cytosolic and Chloroplast Translation Affects Leaf Variegation. Plant Physiol. 2018, 176, 804–818. [Google Scholar] [CrossRef] [Green Version]
- Creff, A.; Sormani, R.; Desnos, T. The Two Arabidopsis RPS6 Genes, Encoding for Cytoplasmic Ribosomal Proteins S6, Are Functionally Equivalent. Plant Mol. Biol. 2010, 73, 533–546. [Google Scholar] [CrossRef]
- Sun, L.; Yu, Y.; Hu, W.; Min, Q.; Kang, H.; Li, Y.; Hong, Y.; Wang, X.; Hong, Y. Ribosomal Protein S6 Kinase1 Coordinates with TOR-Raptor2 to Regulate Thylakoid Membrane Biosynthesis in Rice. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-Y. Molecular Cloning of Low-Temperature-Inducible Ribosomal Proteins from Soybean. J. Exp. Bot. 2004, 55, 1153–1155. [Google Scholar] [CrossRef] [PubMed]
- Romani, I.; Tadini, L.; Rossi, F.; Masiero, S.; Pribil, M.; Jahns, P.; Kater, M.; Leister, D.; Pesaresi, P. Versatile Roles of Arabidopsis Plastid Ribosomal Proteins in Plant Growth and Development: Different Functions of Plastid Ribosome Subunits. Plant J. 2012, 72, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Szick-Miranda, K.; Zanial, A.S.; Zanial, A.S.; Abidayo, S.; Slater, K.L.C. Analysis of RPS15aE, an Isoform of a Plant-Specific Evolutionarily Distinct Ribosomal Protein in Arabidopsis Thaliana, Reveals Its Potential Role as a Growth Regulator. Plant Mol. Biol. Rep. 2010, 28, 239–252. [Google Scholar] [CrossRef]
- Lu, C.; Xie, Z.; Yu, F.; Tian, L.; Hao, X.; Wang, X.; Chen, L.; Li, D. Mitochondrial Ribosomal Protein S9M Is Involved in Male Gametogenesis and Seed Development in Arabidopsis. Plant Biol. J. 2020, 22, 655–667. [Google Scholar] [CrossRef]
- Ma, Z.; Dooner, H.K. A Mutation in the Nuclear-Encoded Plastid Ribosomal Protein S9 Leads to Early Embryo Lethality in Maize. Plant J. 2004, 37, 92–103. [Google Scholar] [CrossRef]
- Qiu, Z.; Chen, D.; He, L.; Zhang, S.; Yang, Z.; Zhang, Y.; Wang, Z.; Ren, D.; Qian, Q.; Guo, L.; et al. The Rice White Green Leaf 2 Gene Causes Defects in Chloroplast Development and Affects the Plastid Ribosomal Protein S9. Rice 2018, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Tzafrir, I.; Pena-Muralla, R.; Dickerman, A.; Berg, M.; Rogers, R.; Hutchens, S.; Sweeney, T.C.; McElver, J.; Aux, G.; Patton, D.; et al. Identification of Genes Required for Embryo Development in Arabidopsis. Plant Physiol. 2004, 135, 1206–1220. [Google Scholar] [CrossRef] [Green Version]
- Schultes, N.P.; Sawers, R.J.H.; Brutnell, T.P.; Krueger, R.W. Maize High Chlorophyll Fluorescent 60 Mutation Is Caused by an Ac Disruption of the Gene Encoding the Chloroplast Ribosomal Small Subunit Protein 17. Plant J. 2000, 21, 317–327. [Google Scholar] [CrossRef]
- Chan, Y.L.; Paz, V.; Olvera, J.; Wool, I.G. The Primary Structure of L37—A Rat Ribosomal Protein with a Zinc Finger-like Motif. Biochem. Biophys. Res. Commun. 1993, 192, 590–596. [Google Scholar] [CrossRef]
- Finkelshtein, A.; Khamesa, H.; Tuan, L.A.; Rabanim, M.; Chamovitz, D.A. Overexpression of the Ribosomal S30 Subunit Leads to Indole-3-carbinol Tolerance in Arabidopsis Thaliana. Plant J. 2021, 105, 668–677. [Google Scholar] [CrossRef]
- Nagaraj, S.; Senthil-Kumar, M.; Ramu, V.S.; Wang, K.; Mysore, K.S. Plant Ribosomal Proteins, RPL12 and RPL19, Play a Role in Nonhost Disease Resistance against Bacterial Pathogens. Front. Plant Sci. 2016, 6, 1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Zhang, C.; Li, Q.; Yang, Q.; Gu, M.; Liu, Q. A Residue Substitution in the Plastid Ribosomal Protein L12/AL1 Produces Defective Plastid Ribosome and Causes Early Seedling Lethality in Rice. Plant Mol. Biol. 2016, 91, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xie, C.; Li, W.; Zhang, R.; Jue, D.; Yang, Q. Expression of a Wild Eggplant Ribosomal Protein L13a in Potato Enhances Resistance to Verticillium Dahliae. Plant Cell Tissue Organ Cult 2013, 115, 329–340. [Google Scholar] [CrossRef]
- Song, J.; Wei, X.; Shao, G.; Sheng, Z.; Chen, D.; Liu, C.; Jiao, G.; Xie, L.; Tang, S.; Hu, P. The Rice Nuclear Gene WLP1 Encoding a Chloroplast Ribosome L13 Protein Is Needed for Chloroplast Development in Rice Grown under Low Temperature Conditions. Plant Mol. Biol. 2014, 84, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Jang, S.; Ryu, S.; Lee, S.; Park, J.; Lee, S.; An, G.; Park, S.K. Mutation of Plastid Ribosomal Protein L13 Results in an Albino Seedling-Lethal Phenotype in Rice. Plant Breed. Biotechnol. 2019, 7, 395–404. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, M.; Day, R.C.; Talbot, M.J.; Ivanova, A.; Ashton, A.R.; Chaudhury, A.M.; Macknight, R.C.; Hrmova, M.; Koltunow, A.M. Developmentally Regulated HEART STOPPER, a Mitochondrially Targeted L18 Ribosomal Protein Gene, Is Required for Cell Division, Differentiation, and Seed Development in Arabidopsis. J. Exp. Bot. 2015, 66, 5867–5880. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.; Chen, D.; Wang, Y.; Sun, Y.; Zhao, J.; Sun, M.; Peng, X. Ribosomal Protein L18aB Is Required for Both Male Gametophyte Function and Embryo Development in Arabidopsis. Sci. Rep. 2016, 6, 31195. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Wada, T.; Yamamoto, K.T.; Okada, K. The Arabidopsis STV1 Protein, Responsible for Translation Reinitiation, Is Required for Auxin-Mediated Gynoecium Patterning. Plant Cell 2005, 17, 2940–2953. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Liu, K.; Zhang, S.; Wang, X.; Rogers, K.; Ren, G.; Zhang, C.; Yu, B. STV1, a Ribosomal Protein, Binds Primary MicroRNA Transcripts to Promote Their Interaction with the Processing Complex in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 1424–1429. [Google Scholar] [CrossRef]
- Park, S.-H.; Chung, M.-S.; Lee, S.; Lee, K.-H.; Kim, C.S. Loss of Ribosomal Protein L24A (RPL24A) Suppresses Proline Accumulation of Arabidopsis Thaliana Ring Zinc Finger 1 (Atrzf1) Mutant in Response to Osmotic Stress. Biochem. Biophys. Res. Commun. 2017, 494, 499–503. [Google Scholar] [CrossRef]
- Szakonyi, D.; Byrne, M.E. Ribosomal Protein L27a Is Required for Growth and Patterning in Arabidopsis Thaliana: Ribosomal Protein-Mediated Patterning. Plant J. 2011, 65, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Zsögön, A.; Szakonyi, D.; Shi, X.; Byrne, M.E. Ribosomal Protein RPL27a Promotes Female Gametophyte Development in a Dose-Dependent Manner. Plant Physiol. 2014, 165, 1133–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oristian, D.S.; Sloofman, L.G.; Zhou, X.; Wang, L.; Farach-Carson, M.C.; Kirn-Safran, C.B. Ribosomal Protein L29/HIP Deficiency Delays Osteogenesis and Increases Fragility of Adult Bone in Mice. J. Orthop. Res. 2009, 27, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aseev, L.V.; Boni, I.V. Extraribosomal Functions of Bacterial Ribosomal Proteins. Mol. Biol. 2011, 45, 739–750. [Google Scholar] [CrossRef]
- Finley, D.; Bartel, B.; Varshavsky, A. The Tails of Ubiquitin Precursors Are Ribosomal Proteins Whose Fusion to Ubiquitin Facilitates Ribosome Biogenesis. Nature 1989, 338, 394–401. [Google Scholar] [CrossRef]
- Chen, J.-G. RACK1 Mediates Multiple Hormone Responsiveness and Developmental Processes in Arabidopsis. J. Exp. Bot. 2006, 57, 2697–2708. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Chen, J.-G. RACK1 Genes Regulate Plant Development with Unequal Genetic Redundancy in Arabidopsis. BMC Plant Biol. 2008, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Li, J.-F.; Niu, Y.; Zhang, X.-C.; Woody, O.Z.; Xiong, Y.; Djonović, S.; Millet, Y.; Bush, J.; McConkey, B.J.; et al. Pathogen-Secreted Proteases Activate a Novel Plant Immune Pathway. Nature 2015, 521, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Kakehi, J.-I.; Kawano, E.; Yoshimoto, K.; Cai, Q.; Imai, A.; Takahashi, T. Mutations in Ribosomal Proteins, RPL4 and RACK1, Suppress the Phenotype of a Thermospermine-Deficient Mutant of Arabidopsis Thaliana. PLoS ONE 2015, 10, e0117309. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MR8G62IP (MTs) | MR1G32IP (non-MTs) | |
|---|---|---|
| Read -1 | 58,784,802.00 | 71,588,702.00 |
| Read -2 | 59,487,917.00 | 73,230,643.00 |
| Read -3 | 59,185,434.00 | 75,556,776.00 |
| Yield (Mbases) | 177,458,153.00 | 220,376,121.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valencia-Lozano, E.; Herrera-Isidrón, L.; Flores-López, J.A.; Recoder-Meléndez, O.S.; Barraza, A.; Cabrera-Ponce, J.L. Solanum tuberosum Microtuber Development under Darkness Unveiled through RNAseq Transcriptomic Analysis. Int. J. Mol. Sci. 2022, 23, 13835. https://doi.org/10.3390/ijms232213835
Valencia-Lozano E, Herrera-Isidrón L, Flores-López JA, Recoder-Meléndez OS, Barraza A, Cabrera-Ponce JL. Solanum tuberosum Microtuber Development under Darkness Unveiled through RNAseq Transcriptomic Analysis. International Journal of Molecular Sciences. 2022; 23(22):13835. https://doi.org/10.3390/ijms232213835
Chicago/Turabian StyleValencia-Lozano, Eliana, Lisset Herrera-Isidrón, Jorge Abraham Flores-López, Osiel Salvador Recoder-Meléndez, Aarón Barraza, and José Luis Cabrera-Ponce. 2022. "Solanum tuberosum Microtuber Development under Darkness Unveiled through RNAseq Transcriptomic Analysis" International Journal of Molecular Sciences 23, no. 22: 13835. https://doi.org/10.3390/ijms232213835